
2
Sustaining and Advancing Ocean Color Research and Operations
Ocean color satellites provide a unique vantage point for observing the changing biology in the surface ocean. Space observations have transformed biological oceanography (Box 2.1) and are critical to advance our knowledge of how such changes affect important elemental cycles, such as the carbon and nitrogen cycles, and how the ocean’s biological processes influence the climate system. In addition, ocean color remote sensing allows scientists to assess changes in primary production, which forms the base of the marine food chain. Thus, continuous satellite observation of ocean color is essential to monitoring the health of the marine ecosystem and its ability to sustain important fisheries, especially in a time of global change. Any interruption in the ocean color record would severely hamper the work of climate scientists, fisheries and coastal resource managers, and an expanding array of other users, from the military to oil spill responders.
It is increasingly recognized that despite the ocean’s vastness, its resilience is finite. The new National Ocean Policy recognizes the many threats to pelagic and coastal marine environments from human activities. Most notably, coral reef environments are degrading because of rising water temperatures, ocean acidification, and other environmental stressors. Overfishing, coupled with environmental variability and habitat loss, threatens many fish stocks. Other human-caused disturbances include chronic beach contamination, hypoxia in estuaries and the coastal ocean, and water quality degradation due to industrial, agricultural, and residential pollution. The 2010 BP oil spill in the Gulf of Mexico is a prime example of a severe anthropogenic impact on the marine environment.
To support the goals and priorities outlined in the National Ocean Policy (CEQ, 2010) and Ocean Research Priorities Plan (JSOST, 2007), continued monitoring of the ocean’s ecosystems on a global scale is essential. The continuity, global coverage, and high temporal and spatial resolution of ocean color products make remote sensing a critical tool for monitoring and characterizing ocean biology and marine ecosystems. Most of the spatial features that are important for marine ecosystems, i.e., ocean fronts, eddies, convergence zones, river plumes, and coastal regions, cannot be brought into full view and studied without satellite observations. Similarly, ocean color products are crucial for making observations frequently enough to study the timing of processes that can have important effects on living marine resources, such as upwelling, harmful algal blooms1 (HAB), seasonal transition, El Niño events, and oil spills.
The use of remotely sensed ocean color products has become ubiquitous in oceanography and marine resource management. This chapter, while not comprehensive, provides examples of the growing array of research and societal applications and related product requirements. In other words, the chapter details the research and resource management questions that ocean color products can help answer. Applications such as coastal marine resource monitoring require near-real time products at fine spatial and temporal scales available from Geostationary Earth Orbit (GEO) satellites (discussed in Appendix D), while others require global, long-term observations provided by traditional sun-synchronous Low Earth Orbit (LEO) satellites. As detailed below, different types of ocean color sensors will be required to meet these diverse demands.
RESEARCH AND SOCIETAL APPLICATIONS OF OCEAN COLOR PRODUCTS
Many basic research and ecosystem management applications of ocean color have long time frames. Users may have to wait weeks or months for data to be processed, and studies can take years before long-term processes and changes can be detected. But ocean color is also critical to users who rely on it for solving societal challenges on a near-real time basis.
____________________________
1 An algal bloom is defined as a rapid accumulation of algal biomass.
Box 2.1
The Unique Vantage Point from Space
“The ability to derive global maps of chlorophyll-a concentration (milligrams per cubic meter) in the upper ocean from ocean color sensors was a groundbreaking achievement for the oceanographic community” (see figure below; NRC, 2008a).
The unique vantage point from space has revolutionized Earth sciences in general. For the first time, scientists have been able to obtain a global synoptic view of the biomass of phytoplankton in the ocean. This unique observing platform has also allowed scientists for the first time to visualize and study dynamic features, such as mesoscale eddies and ocean fronts, and their impact on ocean biology.
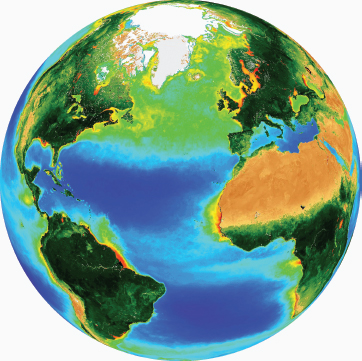
Map of chlorophyll concentration (milligrams per cubic meter) in the upper Atlantic Ocean derived from data obtained by the Sea-viewing Wide Field-of-view Sensor.
SOURCE: SeaWiFS Project, NASA Goddard Space Flight Center, and GeoEye.
Research Applications for Ocean Color Products
Tracking changes in global marine plant biomass is fundamental to ocean biology. Satellite data provide an incredibly powerful tool for observing and quantifying changes in ocean plant biomass over various spatial and temporal scales and for monitoring climate variations and trends. Most of the plant biomass in the ocean comprises microscopic photosynthetic organisms (phytoplankton) that are moved around, mixed, and dispersed by ocean currents and other physical processes. Phytoplankton are different from land plants, which have a large standing biomass that includes large amounts of carbon tied up in structures such as roots and trunks. Phytoplankton standing biomass is comparatively low (about 0.1 percent of land plant biomass) but they grow quickly—in some cases doubling in a single day. Grazing rates on phytoplankton can also be very high. Thus, compared with land plants, phytoplankton biomass is highly variable in space and time and can change much more rapidly in response to changing environmental conditions (Chavez et al., 2011). The dynamic nature of the system, coupled with the difficulty of routinely sampling the global ocean from ships or moorings, demonstrates the incomparable value of remotely sensed ocean color data for tracking phytoplankton biomass variability in different ocean regions over time. Sustaining this record is essential to test many hypotheses about long-term changes in biogeochemical cycles. One example is the current hypothesis that future sea surface warming associated with long-term climate change will result in a decrease in algal biomass, which could affect the ocean’s ability to take up atmospheric CO2 and support marine life that includes valuable fish stocks.
Climate and Biogeochemical Research Applications
How does the ocean’s biology affect the carbon cycle and other biogeochemical cycles?
Carbon is continuously exchanged between the ocean, atmosphere, and land (Figure 2.1). Carbon enters the surface ocean and is dissolved in the water. Compared to the intermediate or deep ocean, the carbon residence time in the surface ocean is relatively short. The biggest reservoir of carbon is in the intermediate and deep ocean. The solubility and biological pumps combined represent the net uptake of carbon by the ocean. When cold, dense water is formed in high latitude, the water containing the dissolved carbon sinks to the deep ocean (solubility pump). Phytoplankton take up some of the carbon in the sun-lit surface layers of the ocean during photosynthesis to produce particulate and dissolved organic carbon. A fraction of this particulate organic carbon can sink (biological pump) to the abyssal plains; an even smaller fraction is buried in the sediments. In these ways, CO2 is removed from surface waters and moved to deep waters and sediments, where it remains for centuries to millennia. It is critical that we continue to improve our understanding of these processes in the ocean, as they are key to the global carbon cycle and other biogeochemical cycles that are essential to life on Earth.
Ocean color observations are important for verification of numerous models that show that the response of ocean biology to changes in ocean circulation may significantly impact the air-sea balance of CO2 in the future (e.g., Sarmiento and Le Quéré, 1996; Sarmiento et al., 1998; Joos et al., 1999; Matear and Hirst, 1999; Plattner et al., 2001; Friedlingstein et al., 2006) and for determining if changes are actually occurring. In addition, ocean color data provide various methods to identify areas of nitrogen fixation (Subramanium et al., 2002; Westberry et al., 2005), which have the potential to alter global biogeochemical cycles.
Ship-based observations provide insufficient spatial coverage to quantify these biogeochemical processes. Because phytoplankton blooms and their associated areas of high productivity are such large-scale yet short-lived phenomena, it is impossible to survey large enough areas of the ocean with ships in order to map phytoplankton productivity and changes in the carbon cycle. Only since the availability of ocean color satellites have scientists been able to routinely estimate global net primary production on weekly to interannual time scales and thus to detect global trends (NRC, 2008a).
Biological uptake of nutrients and carbon at the surface of the ocean plays a crucial role in linking the carbon-rich reservoir of the deep ocean with the atmosphere. Indeed, there are indications that changes in Southern Ocean circulation, together with the response of biology to these changes, may already be affecting the oceanic uptake of CO2 from the atmosphere (Le Quéré et al., 2007; Lovenduski et al., 2007, 2008). The Southern Ocean is estimated to have weakened as a CO2 sink between 1984 and 2004, relative to the trend expected from the corresponding large increase in atmospheric CO2 (Le Quéré et al., 2007). The increase in dissolved CO2 in the surface ocean is also making the ocean more acidic, which has important implications for many marine organisms and ecosystems (Doney et al., 2009; NRC, 2010). Ocean acidification has the potential to reduce rates of calcification, which could lower the efficiency of the biological pump, because de-calcified organisms sink less rapidly and therefore transport organic carbon to the deep sea at a slower rate. These possible effects of ocean acidification are only now beginning to be investigated (Hofmann and Schellnhuber, 2009).
While almost all marine primary production occurs in the sun-lit surface layer of the ocean, large pools of macro-nutrients are found in the deep ocean. In nutrient-limited regions of the surface ocean, vertical upwelling brings nutrients to the surface and drives primary production (Lewis et al., 1986). With some exceptions, nitrate is the nutrient that limits phytoplankton growth in the surface ocean on short time scales (Lewis et al., 1986). Some organisms
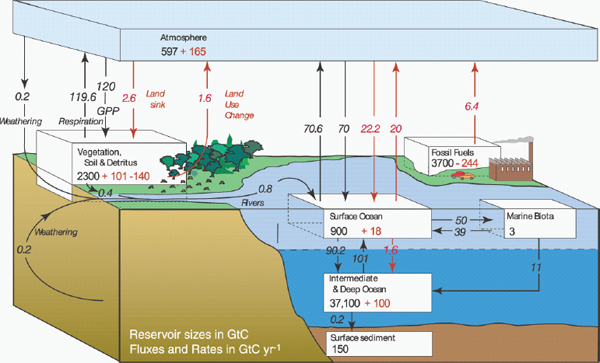
FIGURE 2.1 “The global carbon cycle for the 1990s, showing the main annual fluxes in Gigatons of Carbon (GtC) yr–1: pre-industrial ‘natural’ fluxes in black and ‘anthropogenic’ fluxes in red (modified from Sarmiento and Gruber, 2006, with changes in pool sizes from Sabine et al., 2004). The net terrestrial loss of –39 GtC is inferred from cumulative fossil fuel emissions minus atmospheric increase minus ocean storage. The loss of –140 GtC from the ‘vegetation, soil, and detritus’ compartment represents the cumulative emissions from land use change (Houghton, 2003), and requires a terrestrial biosphere sink of 101 GtC (in Sabine et al., given only as ranges of –140 to –80 GtC and 61 to 141 GtC, respectively; other uncertainties given in their Table 1). Net exchanges of anthropogenic carbon with the atmosphere are based on IPCC WGI Chapter 7. Gross fluxes generally have uncertainties of more than ±20 percent but fractional amounts have been retained to achieve overall balance when including estimates in fractions of GtC yr–1 for riverine transport, weathering, deep ocean burial, etc. ‘GPP’ is annual gross (terrestrial) primary production. Atmospheric carbon content and all cumulative fluxes since 1750 are as of end 1994.”
SOURCE: IPCC, 2007; used with permission from Intergovernmental Panel on Climate Change.
(e.g., Trichodesmium) can thrive in nitrate-depleted waters by fixing dissolved nitrogen. The fixed nitrogen becomes subsequently available to other photosynthetic organisms and might shift the ecosystem toward phosphorus limitation (Karl et al., 1997; Cullen, 1999; Tyrrell, 1999).
Different methods exist to identify nitrogen fixation from satellite ocean color remote sensing (Subramanium et al., 2002; Westberry et al., 2005). For example, large blooms of chlorophyll-containing phytoplankton in the southwest Pacific, observed by both Coastal Zone Color Scanner (CZCS) and the Sea-viewing Wide Field-of-view Sensor (SeaWiFS), have been identified as Trichodesmium blooms (Dupouy et al., 1988, 2000; Westberry and Siegel, 2006). Satellite images also revealed large blooms in late summer in the oligotrophic Pacific northeast of Hawaii (Wilson and Qiu, 2008). Based on biological observations made in situ, those blooms have been attributed to nitrogen fixers or vertically migrating diatom mats (Wilson et al., 2008). The nitrogen provided by nitrogen fixation could contribute substantially to the total available nitrogen for oceanic new production (Capone et al., 1997; Gruber and Sarmiento, 1997; Karl et al., 1997). While total nitrogen fixation is estimated at 110 Tg y–1 (Gruber and Sarmiento, 1997), nitrogen fixation from Trichodesmium species is estimated at 80 Tg y–1 (Capone and Carpenter, 1999). Improving our understanding of the spatial and temporal patterns and production of these blooms is essential because of their size, duration and potential to alter global climate through changes in the biological pump (Michaels et al., 2001; Sañudo-Wilhelmy et al., 2001).
Ocean color data have also revealed extensive open-ocean blooms in the northeast Pacific Ocean (Wilson, 2003; Wilson et al., 2008) and southeast of Madagascar (Longhurst, 2001; Srokosz et al., 2004; Uz, 2007). These blooms are unusual in that they occur in nutrient-depleted regions of the
ocean, and the physical mechanisms that deliver nutrients to support them remain unknown. The discovery highlights how little we know about the open ocean and how our understanding of the complexity of marine ecosystems evolves. Ocean color data have played and will play a crucial role in the discovery process.
How is the global marine phytoplankton biomass changing in response to short-term climate variability and long-term climate trends?
SeaWiFS was launched in August 1997 and contributed to the understanding of the subsequent 1997-1998 El Niño (IOCCG, 2008). The satellite data combined with in situ data from the equatorial Pacific helped scientists understanding El-Niño Southern Oscillation (ENSO) dynamics and their impacts on ecosystems. For example, the deepening of the thermocline and associated weakening of upwelling along the equator and in the coastal ecosystems lowers ocean productivity and causes substantial decreases in the anchovy fisheries of Peru and Chile (Alamo and Bouchon, 1987; Escribano et al., 2004).
Moreover, satellite ocean color products have demonstrated that El Niño effects are not restricted to the equatorial and coastal upwelling regions (IOCCG, 2008). For example, during the 1997-1998 ENSO event, the Transitional Zone Chlorophyll Front, an important region of the North Pacific ecosystem, was shifted about 5°S of its regular position (Bograd et al., 2004). Also, decreased chlorophyll concentrations were observed across most of the subtropical Pacific (Wilson and Adamec, 2001). This basin-scale response, detected with satellite observations, had not been previously detected with in situ observations.
One of the most important challenges for marine science is to distinguish ocean changes caused by interannual variability (e.g., ENSO variability) from trends caused by long-term climate changes or other human effects (Chavez et al., 2011). Recent satellite studies show a general decrease in chlorophyll either in the mid-ocean gyres with some regional variability (Vantrepotte and Melin, 2009) or in the “stratified parts of the low-latitude ocean” (Behrenfeld et al., 2006). The same studies show chlorophyll increases in other parts of the world’s ocean, based on SeaWiFS observations (Antoine et al., 2005; Gregg et al., 2005; Behrenfeld et al., 2006; Polovina et al., 2008; Vantrepotte and Melin, 2009). Sea surface temperature (SST)-based indexes of stratification show a relation between decreasing chlorophyll and NPP and increasing stratification, which suggests a link between reduced phytoplankton abundance and a gradual warming of ocean surface waters (Behrenfeld at al., 2006). Devred et al. (2009) showed that in the Northwest Atlantic, the trends differed among ecological provinces studied.
SeaWiFS was launched during the ENSO event that began in 1997—one of the strongest events of the twentieth century—and thus the initial years of the record are strongly biased by the ENSO effects. Analyses of SeaWiFS data with CZCS observations from 1979 to 1983, and SST observations from both periods, suggest that the basin-scale phytoplankton responses were related to the Pacific Decadal Oscillation and the Atlantic Multidecadal Oscillation with little evidence to suggest long-term trends (Martinez et al., 2009). Comparing satellite records with the results of climate models incorporating ocean ecosystems and biogeochemical cycles indicate that the magnitude of the chlorophyll changes observed during the SeaWiFS era were not unusual. Comparable changes have occurred during multiple-year intervals in the past and can be accounted for by interannual variability (Henson et al., 2010; Yoder et al., 2010).
Because the time-series of remotely sensed phytoplankton chlorophyll is not long enough to detect long-term trends in response to climate change (see Box 2.2), Secchi disk
Box 2.2
How Long Before a Time-Series Can Reveal Long-Term Climate Trends?
Using the methodologies of trend detection, Henson et al. (2010) concluded that 40 years of observations will be required to sort out the effects of natural modes of climate variability, such as ENSO, from trends related to a changing climate and changing ocean. Climate data often contain autocorrelation and large interannual and decadal natural variability that tend to increase the number of years necessary for trend detection (Tiao et al., 1990; Weatherhead et al., 1998). Other factors that make trend detection in climate data challenging include a change in the measurement procedures (e.g., instrument replacement) or a data gap. Changes in measurement procedures can affect data by introducing artificial shifts and/or gradual bias and thus affect the ability to detect trends (Peterson et al., 1998). Such biases can be confused with the magnitude of a real trend and must be corrected for or otherwise taken into account while estimating the trend. Weatherhead et al. (1998) take gaps in the data into account and show that in the worst scenario, the years required to detect a trend can increase by 50 percent.
To arrive at the 40-year time frame, Henson et al. (2010) assume that there will be no interruption in satellite data. This number varies according to the satellite data characteristics and the expected trend magnitude (estimated by using three ocean biogeochemical models) in 14 different regions. If the continuity of measurements is broken over this period, as many as 20 additional years of observations will be necessary. Distinguishing the effects of ocean cycles from long-term trends, and identifying the implications for marine ecosystems, is a major challenge that ocean satellites can help to resolve.
records that date to 1899 have been used as a proxy for phytoplankton chlorophyll (Falkowski and Wilson, 1992; Hou et al., 2007; Boyce et al., 2010). These records have been interpreted as showing an annual decline of about 1 percent in global median chlorophyll (Boyce et al., 2010), which would have major ramifications for the global marine ecosystem. Although the conclusions of Boyce et al. (2010) are challenged by some (Mackas, 2011; McQuatters-Gollop et al., 2011; Rykaczewski and Dunne, 2011), they are consistent with the hypothesis (see also Box 2.3) that increasing ocean warming contributes to changes in the marine ecosystems, which has implications for biogeochemical cycling, fishery yields, and ocean circulation. However, longer records of remotely sensed chlorophyll will be required to detect associated trends in phytoplankton biomass and to improve the estimates of regional rates of change. Assembling those records will be possible only with continuous satellite observations of ocean color. Future research to detect trends in phytoplankton chlorophyll will also have to consider changes in the phytoplankton assemblages and in the relative contribution to the Lw signal from chlorophyll and color dissolved organic matter (CDOM) (Dierssen, 2010). The color of the ocean is driven by the optical properties of the dissolved and particulate materials (including living phytoplankton) in seawater, modulated by the optical properties of pure seawater itself. Thus, the ocean color signals will reflect the combined influences of these materials, all of which vary in abundance, in time, and in spectral characteristics. Detecting these trends and the changes in the relative contribution from CDOM will only be possible with advanced sensor capabilities such as additional spectral bands.
How does ocean color affect radiative heat transfer in the climate system?
To predict the climate accurately, SST has to be simulated correctly. Because older models had such coarse vertical resolution in the upper ocean, variability in the absorption by chlorophyll and CDOM did not make much of a difference in computing depth-dependent heating rates. Recent research has shown that the absorption of light by chlorophyll and CDOM can have a major effect on SST and resulting climate predictions (Marzeion et al., 2005). This discovery demonstrates that the current distribution of ocean color is key to determining the pattern of tropical SSTs. Moreover, as much of the absorbing material is CDOM rather than chlorophyll (Siegel et al., 2005a), these results highlight the importance of understanding CDOM dynamics and how they may change under different climates. There is a rapidly growing recognition of the importance of including these processes in climate models, using primarily satellite-based observations of ocean color for re-interpreting observations, for reanalysis simulations and ecosystem models, and for simulating future conditions.
Studies that examine the role of ocean color in determining large-scale climate changes through altering the absorption profile of solar radiation have yielded somewhat inconsistent results. Some model studies resulted in cooling of the equatorial waters (Nakamoto et al., 2001; Sweeney et al., 2005) while others yielded increased surface heating (Marzeion et al., 2005; Lengaigne et al., 2007) when a realistic distribution of chlorophyll-dependent absorption was included in their models.
Box 2.3
How Might Climate Change Affect the Ocean’s Biology in the Future?
Bopp et al. (2001) carried out the first modeling study to demonstrate how increased stratification and associated processes tend to reduce nutrient supply, and thus biological productivity, in low latitudes. At the same time, these climate-related changes increase the length of the growing season and thus biological productivity in high latitudes, where nutrients are more abundant and less affected by the increased stratification (Bopp et al., 2001). Sarmiento et al. (2004) compared a suite of coupled climate models and illustrated how changes in ocean properties shift the boundaries of biomes in the ocean. More recent studies (e.g., Schneider et al., 2008; Steinacher et al., 2010) compared model simulations that have been used to predict ocean ecosystems. The differences between the models are large; subtle interactions between different drivers of biology and the response of these drivers to climate change yield results that may seem counterintuitive (e.g., Rykaczewski and Dunne, 2010).
Testing of the models with chlorophyll and primary production estimated on the basis of satellite ocean color observations is crucial for developing the models and increasing our confidence in them (Doney et al., 2009), particularly with regard to changes in the seasonal cycle. However, because the chlorophyll: carbon ratio can change substantially in response to changing light levels, it is possible that chlorophyll-based algorithms will not capture relatively small changes in net seasonal productivity. Validation of new products—such as carbon biomass from particulate backscatter (Behrenfeld et al., 2005) or from chlorophyll (Sathyendranath et al., 2009), particle size structure from the backscatter spectrum (Kostadinov et al., 2009), and production associated with individual functional groups (Alvain et al., 2008; Uitz et al., 2010)—is likely to be critical in mechanistically improving the models and in distinguishing changes associated with anthropogenic climate change from those associated with natural variability (Henson et al., 2010).
Recent work suggests that the ambiguity may arise from the fact that different regions respond differently to perturbations in shortwave absorption. In the relatively
stagnant eastern oceanic margins, associated with the oxygen minimum zones, trapping solar radiation closer to the surface cools deeper waters. Because the colder water is essentially trapped in place, it results in cooling the marginal zone (Gnanadesikan and Anderson, 2009). That also results in stabilizing the Walker2 circulation and dampening El Niño (Anderson et al., 2009). In contrast, trapping solar radiation closer to the surface in the gyres results in surface heating, as the deeper, cooler waters tend to be carried away and brought up along the equator. That in turn tends to spin down the Hadley cells, allowing convection to move more freely in the tropics and increasing the amplitude of ENSO. At high latitudes a different scenario is observed, where phytoplankton blooms result in increased cloud cover and cloud albedo, which decreases the local precipitation and incoming solar radiation (Krüger and Graßl, 2011). Ocean color remote sensing will be an important tool to understanding these ocean-climate feedbacks.
How variable in space and time are plant physiology and functional groups?
Some ocean color satellites can measure fluorescence emitted by phytoplankton (Moderate Resolution Imaging Spectroradiometer [MODIS] and Medium-Resolution Imaging Spectrometer [MERIS]) (Gower et al., 2004; Gower and King, 2007; Behrenfeld et al., 2009). This fluorescence measurement can be used to assess phytoplankton physiology, especially as it relates to nutrient availability and irradiance (Kromkamp and Peene, 1995). A global analysis of chlorophyll fluorescence using MODIS data found generally high values associated with waters depleted in iron, and low fluorescence where other environmental factors control growth (Behrenfeld et al., 2009; Sathyendranath et al., 2009). An important component of phytoplankton physiology is the cellular ratio of chlorophyll to carbon (Chl:C). Because changes in light and temperature conditions can impact the Chl:C ratio, observed changes in chlorophyll are not always an accurate proxy for changes in phytoplankton biomass. Satellite ocean color data can be used to derive global fields of the Chl:C ratio, which have strong seasonality and variability driven by light, nutrients, and temperature (Behrenfeld et al., 2005).
Among the big challenges in ocean color research is to move beyond chlorophyll and to identify specific types and functional groups of phytoplankton on the basis of Lw data (e.g., Brewin et al., 2011). Identifying functional groups is important because of their different effects on ecosystem processes, such as their different capabilities for exporting organic matter, including carbon, to depth. Therefore, it is important to understand where the functional groups are found, how they vary interannually, and how they might respond to climate change.
Because different phytoplankton species have different optical properties, they can sometimes be identified with careful analyses of the different wavebands of ocean color. For example, coccolithophores are highly reflective. Under bloom conditions they turn the water a turquoise color that appears as a milky colored patch in satellite images that can be easily discerned by remote sensing. Coccolithophores were the first type of phytoplankton to be specifically isolated by using satellite data (Holligan et al., 1983a; Ackleson et al., 1994; Brown and Yoder, 1994; Brown and Podesta, 1997; Tyrrell, 1999; Gordon et al., 2001; Smyth et al., 2002). There also has been some success in identifying Trichodesmium by using satellite ocean color data (Subramaniam et al., 2001; Westberry et al., 2005; Westberry and Siegel, 2006). Ocean color data also have been used to identify functional groups of phytoplankton, such as haptophytes, Prochlorococcus, cyanobacteria, and diatoms (Sathyendranath et al., 2004; Alvain et al., 2005, Aiken et al., 2007) and to infer the distribution of different size classes of phytoplankton (Ciotti and Bricaud, 2006; Devred et al., 2006; Loisel et al., 2006; Uitz et al., 2006; Hirata et al., 2008; Kostadinov et al., 2010). Increasing the spectral resolution of satellite sensors will improve methods for detecting functional groups from space.
Fisheries and Ecosystem-Based Management
Sustaining the health and resilience of our marine ecosystems is a high national priority. With the creation of the first National Policy for the Stewardship of the Oceans, Our Coasts, and the Great Lakes (CEQ, 2010) it is now the policy of the nation to “protect, maintain, and restore the health and biological diversity of ocean, coastal, and Great Lakes ecosystems and resources” (CEQ, 2010).
The National Oceanic and Atmospheric Administration’s (NOAA) National Marine Fisheries Service (NMFS) is responsible for conserving, protecting, and managing living marine resources in order to maintain a healthy, functional marine ecosystem and the economic opportunities it provides. In this context, NOAA’s fisheries service encompasses commercial fish stocks and all living marine resources (LMR), including threatened and endangered species of fish, marine mammals and invertebrates, and the harvesting and management of commercial fish species. Whereas satellite ocean color products have been used to help harvest fish more efficiently in India and Japan (Wilson et al., 2008; Saitoh et al., 2009), the following section focuses on how ocean color products are used in the United States to assess and manage fisheries and ecosystem health.
____________________________
2 Walker circulation refers to a conceptual model of the air circulation in the tropics. It was first described by Gilbert Walker. According to this model, the air moves along the surface of the Pacific Ocean from east to west, caused by a pressure gradient force that results from a high pressure system over the Eastern Pacific and a low pressure system over Indonesia. The air circulates back at high altitude towards the East, closing the loop of the Walker circulation.
How does environmental variability affect fish stocks and ecosystem health?
Fish stock assessments provide the technical basis for setting annual fish quotas and other management measures to achieve optimum yield while avoiding overfishing and ecosystem harm. At a minimum, a quantitative stock assessment requires monitoring of catch, abundance, and biological characteristics of the stock. Achieving a balance between exploitation and conservation requires substantial information about the stock, its fishery, the ecosystem, and the habitat. However, the environmental factors influencing fish populations are complex, and many remain poorly understood.
Ocean color products are helping to answer a fundamental research question in fisheries oceanography: How does environmental variability affect annual recruitment?3 Because most fish have a planktonic larval stage, successful recruitment depends on the availability of a suitable food source such as phytoplankton. Thus, fish reproduction tends to coincide with the seasonal peak in phytoplankton abundance (IOCCG, 2008). The Cushing-Hjort or match-mismatch hypothesis suggests that recruitment success is related to the match in timing between spawning and the seasonal phytoplankton bloom (Cushing, 1990). Because of the limited spatial and temporal resolution of traditional ship-based measurements, testing that hypothesis requires satellite-based observations. On the Nova Scotia Shelf, for example, highly successful year classes of haddock were associated with exceptionally early spring blooms of phytoplankton, which supports the match-mismatch hypothesis (Platt et al., 2003). Similarly, the timing of the spring bloom and the growth rate of shrimp are correlated (Fuentes-Yaco et al., 2007).
Further, resource managers and scientists recognize the need to move toward ecosystem-based management of fisheries (Browman and Stergiou, 2005; Rosenberg and McLeod, 2005; Sherman et al., 2005; Frid et al., 2006). This provides new impetus to improve our understanding of environmental factors that influence fish stock dynamics and to make environmental variability an integral part of the assessment process. Therefore, routine monitoring of ecosystem parameters such as chlorophyll and marine primary productivity will be increasingly important.
Many spatial features that characterize ecosystems and ecosystem variability—such as ocean fronts, eddies, convergence zones, and river plumes—can be resolved only with satellite data (Holligan et al., 1983b; NRC, 2008a). Moreover, remote sensing of primary productivity and chlorophyll facilitates monitoring the base of the oceanic food chain, which is part of the assessment strategy for large marine ecosystems (Sherman and Hempel, 2008; Chassot et al., 2011; Sherman et al., 2011). For example, satellite chlorophyll data are a key indicator used in the California Current Integrated Ecosystem Assessments (NOAA, 2011).
How to assess the impact of climate change on fisheries?
There is considerable long-term temporal variability in fish stocks. It is a challenge to differentiate the effects of interannual variability and overfishing from long-term changes such as regime shifts (IOCCG, 2008). These regime shifts are characterized by relatively rapid changes in baseline abundances of both exploited and unexploited species (Kendall and Duker, 1998). In addition to SST, changes in chlorophyll can be an index to detect these climate regime shifts early on because these measurements are available from satellites with little time lag and provide basin-scale and global views. To sustain fisheries, management practices need to be flexible enough to recognize and accommodate ecosystem-wide regime shifts (Polovina, 2005).
Long-term changes in ecosystems can be linked to ocean and atmosphere parameters (Mantua et al., 1997; Hare and Mantua, 2000; Peterson and Schwing, 2003). For example, a regional change from a cool to warm climate in the North Pacific in the 1970s coincided with a shift from a shrimp-dominated ecosystem to one dominated by several species of groundfish (Botsford et al., 1997; Anderson and Piatt, 1999). Similar shifts have been observed for many different species and in all ocean basins. Yet the mechanisms that link changes in population abundance to large-scale ocean and atmosphere dynamics are not always clear (Botsford et al., 1997; Baumann, 1998), and the relationships are not always constant (Solow, 2002).
In most cases the relationship between chlorophyll and a specific fish stock is indirect. However, for herbivorous species such as anchovies and sardines, the link can be direct (Ware and Thomson, 2005). Satellite-derived measurements of chlorophyll data provide a fundamental measurement of the base of the oceanic food web that is key to measuring ecosystem changes on a global scale. For example, by extrapolating satellite-derived values of net primary productivity up several trophic levels, Wilson et al. (2009) calculated that the biomass of fish in the ocean are a major but previously unrecognized source of oceanic carbonate that can contribute significantly to the marine inorganic carbon cycle. Satellite measurements of primary productivity also are important for assessing the distribution and diversity of marine organisms (Rosa et al., 2008). In addition, ocean color remote sensing can be used to indirectly extrapolate to benthic habitat or to export production from the base of the euphotic zone (Laws et al., 2000).
Understanding the mechanisms that link changes in fish population to the climate system will require long-term time-series of key ecosystem parameters. Such understanding will be central to developing new management strategies, given the projected long-term climate trends. Modeling studies suggest that climate change will result in a large-scale redis-
____________________________
3 Annual recruitment definition: the number of new individuals of a stock that enter the fishery in a given year.
tribution of fisheries’ catch potential (Cheung et al., 2008b). The high-latitude regions are projected to experience a 30 to 70 percent increase in catch potential, while a decline of up to 40 percent is projected for low-latitude regions. These major changes will be accompanied by changes in biodiversity (Cheung et al., 2008a).
Global primary production measurements from satellites, together with fish catch statistics and food web models, can be used to estimate the carrying capacity of the world’s fisheries (IOCCG, 2008; Chassot et al., 2010). Much of the world’s fish catch comes from coastal areas, which are near or beyond their carrying capacity based on recent estimates (Pauly and Christensen, 1995). Ocean color data, therefore, is paramount for managing fisheries over the long term.
How to characterize ocean habitats remotely?
Reflectance of the seafloor holds great promise for characterizing and monitoring shallow benthic habitat from space. In “optically shallow waters,” where the reflectance of the seafloor influences the ocean color observed at the sea surface, ocean color is used to characterize bottom types. Ocean color remote sensing offers a cost-effective and repeatable approach for mapping and potentially quantifying benthic substrate. In addition, ocean color can help detect large-scale changes in the health of coastal pelagic and benthic ecosystems, particularly in large homogenous regions (Dekker et al., 2006). Current and planned U.S. research satellites, however, do not provide the required high spatial and spectral resolution for all applications in this class. Many resource managers and other users will likely need a better mix of satellites with superior resolution.
Airborne ocean color sensors have been used to document resilience of benthic features to hurricanes and other large-scale disturbances. Seagrass distributions observed in the eastern portion of the Bahamas Banks near Lee Stocking Island using high-resolution imagery from the Portable Hyperspectral Imager for Low Light Spectroscopy (PHYLLS) were analyzed. Meadows varied from sparse to dense over meter scales (Dierssen et al., 2003). Although Hurricane Floyd inflicted significant damage to structures on the adjacent island, ocean color data revealed that turtle-grass distributions in the region were virtually undisturbed. Satellite observations from Landsat and other multispectral imagers have been used to determine the space-time distribution of Giant Kelp biomass in California and its relationship to nutrient availability and surface wave disturbance (Cavanaugh et al., 2011).
In addition, ocean color imagery can easily identify turbidity or sediment re-suspension events, often caused by storms or high winds, because of the high backscattering signals of the suspended sediments (Acker et al., 2004; Chen et al., 2007; Hu and Müller-Karger, 2007; Dierssen et al., 2009). For example, after the passage of Hurricane Dennis in July 2005, substantial sediment re-suspension covered nearly the entire west Florida shelf. Such turbidity events can affect seagrasses or corals in two ways: these events can be associated with potential eutrophication, which can lead to hypoxic conditions that increase benthic mortality, and increases in water column turbidity that reduces available light for photosynthesis (Adjeroud et al., 2002; Zimmerman, 2006).
How to manage protected species?
NOAA’s Office of Protected Resources manages approximately 300 species, including those listed under the Endangered Species Act and the Marine Mammal Protection Act. The Endangered Species Act requires NOAA to designate “critical habitat” and to develop and implement recovery plans for threatened and endangered species. As the only satellite measurement of the marine ecosystem, ocean color data are a crucial tool for characterizing the habitat and behavior of protected species. The Transition Zone Chlorophyll Front (TZCF), which migrates seasonally between 30°N and 40°N in the North Pacific, is an important foraging and migration corridor for a number of marine species (Polovina et al., 2001, 2004), including the endangered monk seal. The interannual variability of the TZCF also affects the ecosystem as far south as the northern atolls of the Hawaiian Archipelago—the Kure, Midway, and Laysan Atolls. In some years, the TZCF remains north of those atolls all year. In other years, the TZCF shifts far enough south during winter to infuse the atolls with higher-chlorophyll water, making the region more biologically productive. This variability has been observed to affect the population of the monk seal. After a winter during which the TZCF shifted south, monk seal pup survival increased (Baker et al., 2007).
Ocean color data also have been used to help with the management of the North Atlantic Right Whale, one of the most endangered whale populations with fewer than 400 individuals left (International Whaling Commission, 1998; Kraus et al., 2005). High mortality, especially as a result of ship strikes and whales becoming entangled in fishing gear, limits the recovery of this population. Because whale habitat overlaps with lucrative fishing grounds and shipping lanes of major U.S. ports, reducing mortality is politically and economically challenging (International Whaling Commission, 1998; Kraus et al., 2005). The current management strategy limits adverse impacts by requiring modifications of fishing gear or vessel speeds in regions where and during times when whales are likely to be present. Satellite ocean color data have been used to identify whale feeding grounds. For example, an effort to provide managers with a forecast of right whale distributions has the goal of avoiding whale strikes by merchant ships (Pershing et al., 2009a,b).
Lastly, satellite ocean color data are crucial in biologging studies, in which animals are tagged to obtain measurements as they move undisturbed through their environments. Recent advances in biologging technology have advanced our understanding of the ecology of top predators and have
permitted observations previously unavailable from standard measurement techniques (Bograd et al., 2010). Satellite data make it possible to place the tagging data in an environmental context so that we can understand the foraging and migration patterns. This approach has been used to characterize the behavior and habitat of a wide variety of tagged species, including turtles, penguins, seals, salmon, whales, and sea birds (Hinke et al., 2005; Ream et al., 2005; Polovina et al., 2006; Weng et al., 2007; Block et al., 2011).
Near-Real Time Applications for Ocean Color Products
Applications for ocean color products that require near-real time (NRT) turnaround, from image acquisition until final products reach users, present significant challenges. The best possible analysis must be done with whatever imagery is available at a given moment, which often is not the best imagery for the task. GEO satellites are ideal for collecting NRT data, but the only ocean color geostationary satellite is the Korean Communications Oceanography and Meteorology Satellite (COMS) that currently provides data over the Korean Pacific Ocean. Other approaches are possible; Appendix D describes one cost-effective option to obtain GEO ocean color data.
Military Applications
Much U.S. Navy-sponsored research in the past 10 to 15 years has been directed at coastal and optically shallow waters. The main goal is to characterize the optical properties of the environment for operation of ships, submarines, and divers in the water. A major thrust of the research has been to develop and test airborne hyperspectral sensors and algorithms for mapping bathymetry and bottom type at meter spatial scales in optically shallow waters.4 Recent projects funded by the Office of Naval Research that investigate these and related topics include Coastal Benthic Optical Properties (1997 to 2002) and Hyperspectral Coastal Ocean Dynamics Experiment (1999 to 2004). Other countries, especially Australia and the People’s Republic of China, have invested heavily in the development of airborne hyperspectral imagery capabilities for their coastal waters.
The Navy also uses ocean color to determine water clarity, in order to decide whether particular sensors (such as mine-finding or bathymetric LIght Detection and Ranging sensors [LIDAR]) can be deployed or visibility is adequate for diver operations. The military uses surface measurements of ocean color products to initialize or validate physical-biological ecosystem models, which permit the extrapolation to greater depth of properties such as chlorophyll, absorption, and backscatter that affect visibility and mixed-layer thermodynamics.
Monitoring of Oil Spills
The Oil Spill Response Research Program in the U.S. Department of the Interior pursues techniques for early detection, containment, and clean up of oil spills. Remote sensing can be used to detect spills because the oil changes surface reflectance properties and the color of the ocean. Application of MODIS 250-m imagery can help locate natural oil seeps and improve estimates of natural oil seep rates in the ocean because of the near-daily revisit and wide sun-glint coverage (sensors are usually designed to avoid sun glint, which interferes with obtaining good chlorophyll data, but sun glint is particularly effective at capturing the presence of oil at the surface; Hu et al., 2009). In addition, data from both the MODIS and MERIS sensors were used to track the massive Deep Water Horizon oil spill in the Gulf of Mexico in spring and summer 2010. Surface manifestation of oiled water within the region of sun glint in the full resolution “true color” images was readily apparent. Scientists mapped the extent of the spill and the location of areas of convergence in order to direct ship activities focused on rescuing oiled turtles (Dave Foley, personal communication).
Airborne Visible InfraRed Imaging Spectrometer (AVIRIS) provides full solar spectral reflectance observations that were used during the Deepwater Horizon event to determine the spatial distribution of oil layer thickness. These observations are useful in routine oil spill monitoring and response efforts to differentiate recoverable oil films (> 0.1 mm) from thinner films. The use of multispectral UV-visible detectors, coupled to thermal infrared detectors, allows detection and discrimination of oil films. During spill events, decision makers need access to imagery within two hours of data collection.
However, current satellites are limited in their capacity to assist in oil spill detection. Coarse spatial and temporal resolution, limited spectral bands, cloud-cover issues and the need to operate in conditions of high sunlight have generally restricted the usefulness of ocean color data from low Earth orbit satellites for oil-spill detection (Fingas and Brown, 1997, 2000; Hu et al., 2003). Moreover, current processing methods may not allow for data availability within two hours of data capture. The AVIRIS sampling covered only a small part of the Gulf of Mexico region.
The spatial, temporal, and spectral resolution needed for oil spill recovery planning requires high resolution, hyperspectral ocean color radiometers deployed in geostationary orbit. Geostationary sensors can scan large regions of the ocean over long periods—a clear advantage over airborne sensors. This reduces problems with cloud cover, and the wide coverage can help optimize the deployment of airborne and marine assets. Plans for the proposed hyperspectral ocean color sensor on Geostationary Coastal and Air Pollu-
____________________________
4 Shallow waters are defined as waters where bottom reflectance significantly affects the water-leaving radiance. Depending on inherent optical properties of the water, this depth ranges from less than one to tens of meters.
tion Events mission (GEOCAPE; a decadal survey mission; [NRC, 2007]) call for a spatial resolution of about 300 m and the operational flexibility to observe special events, such as oil spills. In addition, Appendix D describes a cost-effective means to provide ocean color data from commercial GEO satellites, to be demonstrated in fall 2011 by the Commercially Hosted Infrared Payload (CHIRP) program.
Detection and Early Warning of Harmful Algal Blooms
Ocean color products are an important component in forecasting HABs, which requires frequent observations over a large area to assess bloom location and movements (IOCCG, 2008). HABs can sicken or kill animals and humans by degrading the water quality or by producing algal toxins, which harm by direct exposure or through consumption of contaminated food.
For example, some HAB species cause areas of extremely low-oxygen water, which can kill a large portion of the sessile fauna (Rabalais et al., 2001). Other HAB species have caused large kills of birds and marine mammals along the U.S. West Coast (Trainer et al., 2000). To avoid human deaths, such as in eastern Canada in 1987 (Subba Rao et al., 1988; Bates et al., 1989; Wright et al., 1989), fisheries are closed in affected areas, and their closure is associated with a loss in revenue. Toxins released to the atmosphere by Karenia brevis blooms have been linked to respiratory illnesses along the Gulf of Mexico coast (Hoagland et al., 2009).
HAB events adversely affect commercial and recreational fishing, tourism, and valued habitats. Advanced warnings of HABs and estimations of their spatial distributions increase the options for managing these events, avoiding exposure of humans to the toxins, and minimizing the negative effects on local economies and the livelihood of coastal residents.
Various methods are being developed to improve the detection of HABs with ocean color sensors (Stumpf et al., 2003; Hu et al., 2005; Tomlinson et al., 2009; Zhao et al., 2010). However, many challenges remain. In particular, the spatial and spectral resolution of current ocean color sensors can be too coarse to detect features in many coastal regions. In the case of some harmful algal blooms, there may not be any feature among their optical properties that can be used to distinguish them from non-toxic blooms (Sathyendranath et al., 1997). Despite the limitations, satellite ocean color can be an effective tool for monitoring HABs, which NOAA has done in the United States since 2006, producing HAB bulletins twice a week for the Gulf of Mexico (Stumpf et al., 2009). Efforts also are under way to develop ocean color-based operational HAB forecasts in Europe (Johannessen et al., 2006) and Australia (Roelfsema et al., 2006). Recently methods have been developed for predicting domoic acid-producing Pseudonitschia blooms that use satellite ocean color and regional ocean circulation model data products (Anderson et al., 2011). As with monitoring for oil spills, geostationary ocean color satellites (Appendix D) would be invaluable assets for monitoring HABs because they provide the required frequency and spatial resolution to be effective.
Locating Productive Fishing Areas
As fish stocks dwindle, the fishing industry increasingly relies on technology to locate and catch fish quickly and at less cost. Satellite maps of sea surface temperature and ocean color can help increase efficiency by identifying sites of fish aggregation and migration, such as temperature fronts, meanders, eddies, rings, and upwelling areas (Laurs et al., 1984; Fiedler and Bernard, 1987; Chen et al., 2005). In addition, ocean color or temperature gradients can be used to indicate biologically productive regions. Fishermen with knowledge of particular fish species’ temperature ranges and preferences have been using SST from the NOAA polar orbiting satellite for the past 20 years.
However, for ocean color to be of practical use to the fishing industry on a large scale, remotely sensed chlorophyll maps must be made available in near-real time. In Japan and India the national fisheries agencies use ocean color to increase the efficiency of their fishing fleets. For example, data from the Indian Ocean color satellite is used to provide maps of potential fishing zones in NRT (Nayak et al., 2010). In contrast, NMFS does not distribute “fish-finding maps” or provide other services that would compete with commercial interests. Chlorophyll data from the privately owned SeaWiFS satellite were only available on a real-time basis to commercial subscribers. Only clients of the service company can receive custom-tailored maps of ocean color and other satellite-derived data directly onboard their fishing vessels.
Identifying Areas with Potential for Marine Debris Convergence
Marine debris and abandoned fishing nets, also called “ghost nets,” pose a serious hazard to many marine mammals and sea birds (Jacobsen et al., 2010). Endangered sea turtles, seals, and whales are among the species that become entangled in the nets and die. The nets become ensnared on coral reefs and damage the reef structure, destroy flora and fauna that depend on a healthy reef ecosystem (Donohue et al., 2001), and can harm commercial fisheries (Kaiser et al., 1996; Gilardi et al., 2010).
Satellite ocean color data are part of the methods being developed to identify and prioritize the likely locations of marine debris in order to remove it from the ocean. In one instance, satellite ocean color data were used in the subtropical North Pacific to detect probable locations of debris convergence. A field program validated the detection method (Pichel et al., 2007). Satellite measurements of chlorophyll
and the chlorophyll gradient were analyzed in conjunction with observer sightings to generate a map of the likelihood and density of debris. Ocean color data were vital to this identification method; initial efforts using only satellite temperature data were much less effective.
Cruise Support
Near-real time satellite data are invaluable for guiding oceanographic research and operational fishery survey cruises and are used in this manner by NOAA, the Coast Guard, and the research community. The Warm Core Rings experiment in the early 1980s, using the Coastal Zone Scanner (Brown et al., 1985), demonstrated the utility of NRT ocean color data for guiding oceanographic expeditions. NRT data are important for locating blooms, fronts, eddies, and other relevant features for sampling or process studies. The spatial coverage provided by satellite data is crucial for placing ship-based observations in a larger geographic context.
Future Applications
Chapter 5 describes how future enhancements could increase the value of ocean color products. Advances in satellite sensor technology, increased spectral and spatial resolution, and the addition of sensors in geostationary orbits that would allow frequent sampling and even greater spatial resolution would expand the value of ocean color products to climate scientists, the ocean research community and the military, among others (NASA, 2006). Such advances would allow better characterization of coastal environments from space, including the benthic and estuarine environments. LIDAR sensors would expand ocean color observations to greater depths, dramatically enhancing the accuracy of global plant biomass and carbon estimates. Lastly, the Navy is eager to obtain higher spatial and temporal resolution in shallow coastal waters, available from GEO (Appendix D), and would benefit from the vertical resolution provided by LIDAR sensors.
OCEAN COLOR DATA SPECIFICATIONS IN SUPPORT OF OCEAN COLOR APPLICATIONS
As the user community broadens, ocean color product requirements grow increasingly diverse. For example, global biogeochemical modeling studies require global images of relatively coarse spatial resolution, whereas HAB work requires frequent, high-resolution observations only available from GEO (Appendix D).
There are three primary product characteristics for applications of ocean color data: spatial resolution, wavelength resolution, and repeat time. Spatial resolution refers to the area of the ocean surface that corresponds to a single image pixel (i.e., ground sample distance). This can range from less than 1×1 m to more than 1×1 km. Wavelength resolution refers to the number of wavelengths measured and their bandwidths. It can vary from monochromatic (a gray scale with perhaps all visible wavelengths sensed) to multispectral (typically 5-15 wavelengths with each band 10-20 nm wide), to hyperspectral (~30 to a few hundred wavelengths with bandwidths of 10 nm or better). Repeat time refers to the time between successive images of a given location on Earth. It varies from multiple images per day, available only from GEO satellites, to several days for satellites in polar orbit, or on a monthly or less frequent basis (especially in areas with frequent clouds).
TABLE 2.1 Four Types of Satellite Sensors Required to Meet the Observational Needs
| Type | Description | Examples |
| 1 | Polar orbiting sensors with relatively low spatial resolution (1 km) with 8 (or many more) wave bands. | SeaWiFS, MODIS, VIIRS, PACE/ACE |
| 2 | Polar orbiting sensors with medium spatial resolution (250-300 m) and more bands to provide a global synoptic view at the same time as allowing for better performance in coastal waters (but with longer repeat times for global coverage). | MERIS, OCM-1, OCM-2 |
| 3 | Hyper-spectral sensors with high spatial resolution (~30-100 m) in polar orbit and even longer repeat times. | HyspIRI |
| 4 | Hyper- or multi-spectral sensors with high spatial resolution in geostationary orbit. | GOCI, GOCI-II, GEOCAPE |
While a polar orbiting satellite sensor like SeaWiFS can deliver a global image about every three days, it cannot increase its spatial or temporal resolution to respond to requirements for higher resolution during an event such as an oil spill. In such a case, a geostationary satellite (Appendix D) that could increase its sampling frequency for a given location would be ideal. The committee identified four general types of sensors to cover the spectrum of observational needs (Table 2.1).
The required data specifications were determined for each application of ocean color data described in this chapter (Table 2.2).
As Table 2.1 illustrates, the needs of the research and operational community are too diverse to be met by a single satellite. For example, a sensor in polar orbit designed to provide global synoptic coverage will not be able to provide the high spatial and temporal resolution required to meet the operational needs of the military or to be of use during an oil spill response.
TABLE 2.2 Current Applications of Ocean Color Data and Required Data Specifications
| Application | Spatial Resolutiona | Wavelength Resolution | Repeat Timeb | Coverage | Sensor Type | References |
| Research and Societal Applications: | ||||||
| Chlorophyll variability and trends at regional to global scales | 1 km | Multispectral | 2-3 days | Global | 1 | Many |
| Trends and variability in carbon fixation at regional to global scales | 1 km | Multispectral | 2-3 days | Global | 1 or 2 | Many |
| Measure inherent optical properties | 1 km | Multispectral | 2-3 days | Global | 1 | IOCCG, 2006 |
| Phytoplankton physiology; C:chl ratios; physiological states and growth rates | 1 km | Multispectral including fluorescence wave band | 2-3 days | Global | 1 | Behrenfeld et al., 2005, 2009 Sathyendranath et al., 2009 |
| Phytoplankton phenology; Time-series of chlorophyll, primary production, phytoplankton functional types | 4-10 km for open ocean; better for coastal waters | Week-monthly | Global | 1 | Platt and Sathyendranath, 2008 Siegel et al., 2002 | |
| Carbon inventory of the ocean (colored dissolved organic matter, particulate carbon, particulate inorganic carbon, phytoplankton carbon) | 4-10 km for open ocean; better for coastal waters | Often derived from chlorophyll and IOPs | Week-monthly | Global | 1, 2 or 4 depending on application | Balch et al., 2005 Behrenfeld et al., 2005 Gordon et al., 2001 Siegel et al., 2002 |
| Climate change impacts on ocean ecosystem | 1 km | Multispectral | 2-3 days | Global | 1 | Henson et al., 2010 |
| Detection of phytoplankton functional groups | 1 km | Multispectral, selected narrow bands; sometimes derived from IOPs or chlorophyll | Weekly | Global | 1 and 2 | Bracher et al., 2009 Sathyendranath et al., 2004 Alvain et al., 2005 Nair et al., 2008 |
| Heat budget and upper ocean dynamics; air-sea interactions (Diffuse attenuation coefficient for visible solar energy) | 1 km or better | Multispectral | 1 day | Global | 1 | Many papers; e.g., Gnanadesikan et al., 2010 Murtugudde et al., 2002 Ohlmann et al., 1996 Sathyendranath et al., 1991 |
| Fisheries and Ecosystem Based Management: | ||||||
| Ecosystem Based Management (Fisheries) | 1 km | Multispectral | 2-3 days | Global | 1 | |
| Mapping the boundaries of ecological provinces in the ocean and their movement | 4-10 km for open ocean; better for coastal applications | Derived from chlorophyll, SST and other satellite-derived products | Various time scales of interest, from 1 week to 1 year | Global | 1 | IOCCG Report No. 8, 2009 |
| Coastal dynamics (suspended sediment load, sediment transport, river plumes, etc.) | 100 m to 1 km | Multispectral | Hourly to 3 days depending on region and need; | Coastal and estuarine waters | 1, 2, 3, or 4 | Many papers (e.g., Warrick et al., 2004) |
| Application | Spatial Resolutiona | Wavelength Resolution | Repeat Timeb | Coverage | Sensor Type | References |
| Monitor coral reefs | 10 m best, 50 m OK for mapping entire reef systems | Multispectral | Annual | Coastal, 30 N to 30 S, so Geostationary OK | 3 | Hochberg, 2011 Lubin et al., 2001 |
| Monitor sea grass beds and kelp forests and biomass | 3-30 m for biomass (100 m for mapping) | Multispectral | Twice a year | Coastal and estuarine, 60 N to 60 S | 3 | Cavanaugh et al., 2011 Hill and Zimmerman, 2010 |
| Near-Real Time Applications: | ||||||
| Naval application for shallow water bathymetry and bottom classification | 1-10 m | Hyperspectral best, multispectral is still useful | On demand | Coastal waters, as needed | 1-4 | Dekker et al., in press Mobley et al., 2005 |
| Monitoring oil spills | 100 m | Multispectral | Hourly | Episodic regional events | 4 | Hu et al., 2009 |
| Detection of HABs | 100 m to 1 km | Multi, but hyper better | 1 day to monitor blooms | Coastal and estuarine waters | 1 and 2 | Ruddick et al., 2008 |
a Minimum to sustain current capabilities, not necessarily optimal.
b Equatorial revisit.
The extensive list of research and societal applications presented in this chapter demonstrates that ocean color is fundamental to and irreplaceable for a wide array of applications at local to global spatial scales and near-real time to decadal time scales.
Exploring the full potential of ocean color research will require more than sustaining current efforts including major advances in sensor capabilities, atmospheric corrections, and algorithm and product development as further described in Chapter 5. Ocean color has been recognized as an essential climate variable5 by Global Climate Observing System6 (GCOS). To detect long-term climate trends in marine phytoplankton abundance, long time-series of sufficient quality are required, making it imperative that we maintain and advance satellite capabilities. With the anticipated impacts of climate change on the marine ecosystem, monitoring the changes will be essential to managing the diminishing resources in the ocean.
Further, the expanding use of ocean color products in research and resource management demands a range of spatial and temporal product specifications that are beyond the ability of a single satellite mission to deliver (see Chapters 4 and 5 for details). Wavelength resolution requirements range from only a few bands (to determine chlorophyll) to hyperspectral data (for coastal and naval applications as well as for advanced algorithms for atmospheric correction and the separation of phytoplankton absorption from CDOM; see Chapter 3). A polar orbiting satellite can provide a global image about every three days at relative coarse spatial resolution and meet requirements for applications that need a global synoptic view, such as climate research. The same satellite, however, cannot deliver multiple images per day at the high spatial resolution required by the Navy or by an oil spill response. Those scenarios require a GEO satellite (Appendix D).
Conclusion: A mix of orbits and sensors are required to meet the indisputable demand for a continuous ocean color record that will help us to understand changes in the global climate system, assess the health of the marine ecosystem, and sustain important fisheries, among other crucial societal tasks.
____________________________
5 See http://www.wmo.int/pages/prog/gcos/index.php?name=Essential ClimateVariables; accessed June 10, 2010.
6 See http://www.wmo.int/pages/prog/gcos/Publications/gcos-138.pdf.














