
3
Stressors:
Environmental Factors and Their
Effects on the Bay-Delta Ecosystem
Many environmental factors, including water diversions, affect the structure and functioning of biotic communities in the delta. Although it would be convenient if one or only a few of these factors could be identified as the source of the “problem,” or even ranked with some certainty, it is not possible to do that, for at least three reasons: the “problem” is not easily definable, the suite of stressors is complex and interactive, and the ecosystem and its components do not react to any stressor as a single unit.
“The Problem” of the Delta Is Not a Single, Easily Definable Problem
Although the ecosystem has been radically altered over the past 150 years, it nonetheless remains a biologically diverse and productive ecosystem. Some species have thrived, but others, including some listed as threatened or endangered under the federal Endangered Species Act and California’s Endangered Species Act, have declined dramatically. In addition, species composition and environmental conditions in the delta have undergone large changes over the period. Therefore, while an immediate difficulty for some is that concern over some listed species has affected water diversions, “the problem” is harder to define biologically, and is perceived differently by various stakeholder groups, institutions, and other interests.
The Suite of Stressors Affecting Water Quality, Habitat, and Sustainability of the San Francisco Bay Delta Is Complex and Interactive
Interactions among stressors and between stressors and ecosystem processes are common. Nutrient enrichment, toxic chemicals, and temperature, for example, are affected by hydrology and hydrodynamics, that is, the way tides and freshwater flow interact to determine the temporal and spatial variability of the physical environment of the estuary. This complicates the interpretation and evaluation as to positive, negative, or neutral overall effects of any single stressor on the ecosystem and its attributes. Furthermore, species differ in their individual responses to most types of stress. The result is a complex biological, spatial, and temporal mosaic of impacts from this combination of influences.
To some extent, the evaluation of the impacts of these effects also depends on which ecosystem services and needs are of interest or concern, for example, safe and usable water supplies, recreational and commercial fisheries, habitat condition, or public use of the delta. Thus, while it is politically attractive to attempt to rank stressors so as to prioritize societal investments in their amelioration, that task is much more complex than it might at first seem. To some degree, priorities can be defined if the stress, species, place, and time are first prioritized or defined. The stressors discussed below and shown in Figure 3-1 are highly dynamic; that is, they can quantitatively change in time and space depending on changes in human activities (including future management actions), climate, and combinations thereof.
The Ecosystem and Its Components Do Not Necessarily Respond as a Single Unit to Most Environmental Factors
For example, Chinook salmon (Oncorhynchus tshawytscha) spend several years at sea and then return to pass through the delta as adults to spawn; their eggs and young spend time in delta tributaries before passing through the delta on their way to the ocean to mature. Returning adult Chinook salmon always die after spawning, and so they are not susceptible to chronic environmental factors, because they die before such factors can affect them. They also are strong swimmers and therefore most changes in flow patterns in the delta are reasonably small challenges for them. The eggs and young are susceptible to conditions in the tributaries and are exposed to them for considerable periods, and the outmigrating smolts are not as strong swimmers as are the returning adults, and so probably are more susceptible to changes in flow patterns. By contrast, delta smelt (Hypomesus transpacificus) spend their entire (short) lives in the delta and so they can be chronically exposed to contaminants in the water; being smaller and
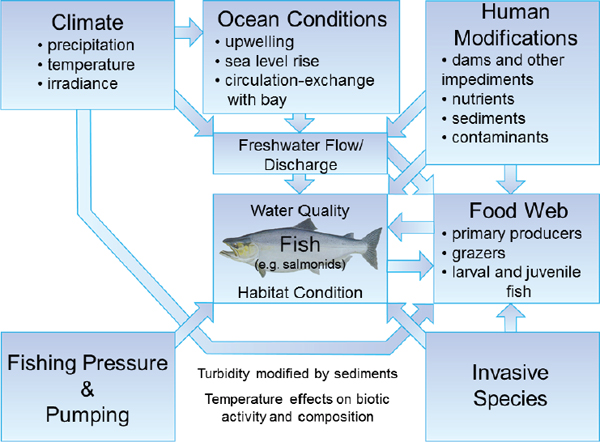
FIGURE 3-1 Conceptual diagram showing the interactive stressors affecting San Francisco Bay Delta water quality, habitat condition, and overall ecosystem structure and functioning. While this figure is focused on key fish species (e.g., salmonids), these are intimately linked to other biotic components of the ecosystem, including planktonic and benthic primary producers, grazers, larval, and juvenile and mature invertebrate and fish species.
SOURCE: Courtesy of A. Joyner, University of North Carolina.
weaker swimmers than even salmon smolts, they likely are more susceptible to changes in flow than salmon are. In addition, the behaviors, food, distribution in the water column, and physiologies of salmon and smelt are different, so even if they are exposed for a time to the same adverse environmental conditions, their responses to them almost certainly are different.
The above discussion compared only two species, but other species are important as well, including those that are not listed. Other biotic components, ranging from phytoplankton to fish, are part of the ecological community and yet they, too, differ in behavior, distribution, physiology, and susceptibility to a wide variety of environmental conditions, including contaminants. Thus most attempts to identify and rank single environmental factors as stressors are very likely to fail, unless the factors can be specifically related to a particular aspect of a species’ life history. Even such
factors as dams, which would appear at first glance to adversely affect only or mainly migratory species like salmon, steelhead (Oncorhynchus mykiss), and green sturgeon (Acipenser medirostris), also affect flow patterns, water temperature and quality, food availability, and so on, and they differentially affect many species, even those that do not migrate. There is a complex interplay between key water quality, habitat, and sustainability issues and the drivers affecting them. Furthermore, uncertainties and scientific gaps exist that further compound the problem (Table 3-1). Indeed, the delta problem is a “wicked” problem in the sense of Rittel and Webber (1973) and Conklin (2005): the problem is hard to define objectively and the nature of the problem depends on the values of those who define it.
For all the above reasons, the committee concludes that only a synthetic, integrated, analytical approach to understanding the effects of suites of environmental factors on the ecosystem and its components is likely to provide important and useful insights that can lead to enhancement of the delta ecosystem and its species.
Although the committee recommends a synthetic, integrated approach to assessing environmental factors, such an approach first requires a description of the individual factors separately. Therefore, we provide such descriptions, covering a variety of environmental factors that are important or potentially important in the following sections. The current set of stressors discussed is not an exhaustive list; rather, they are the most prominent stressors in the delta system in the committee’s judgment. Following this, the committee provides its assessment of each stressor individually.
Physical Environment: Geomorphology and Delta Geometry
Changes in geomorphology of the delta in the last 150 years have been dramatic. Alteration of tidal channels and drainage of wetlands within the delta began for agricultural purposes, but eventually, as new centers of commerce and shipping developed, the drained lands supported urban development. Levees surrounding delta islands isolate most land in the delta from tidal or riverine flooding. Historically, periodic flooding of floodplains and wetlands provided habitat for many species and reduced the risk of downstream flooding. The delta absorbed flood flows to become a vast shallow lake. At its greatest extent prior to the transition to agriculture, the delta covered 1,931 square miles of tidally influenced open water, intertidal flat, and marsh. By 1930, however, 35 percent of the delta had been converted (Thompson 1957), leading a trend of land conversion that established the channel geometry and variability that is present today.
TABLE 3-1 Examples of the Interplay Among Ecosystem Processes (Drivers), Stressors, Science Needs, and Policy
| Driversa | Stressors | Water Policy Issues | Uncertainties and Science Needs |
| Anthropogenic infrastructure changes resulting in changes in freshwater flow and turbidity | Canals. Removing more water from the system. Reservoirs. | Effects: benefits vs. adverse implications for ecosystems | Predicting influences of new water routing? Implications of population growth, water use or conservation? Impediments and benefits to fish passage. |
| Climate change | Temperature: Changing ocean conditions. Changing hydrology. | Will future habitats be suitable for species of concern? Can we save and manage sensitive species? | Can we manage habitats to create refuges and sustain optimal carbon, nutrient, and oxygen cycling? |
| Exports | Entrainment. Indirect effects on hydrodynamics. Nutrient and carbon loadings. Upstream diversions. | How to balance supply reliability with ecosystem requirements. | Effects on fish populations vs. individuals? Quantifying indirect effects? Quantifying effects of upstream diversions? |
| Food quality | Nutrients: N,P,C. Flow. Grazing. | Declining quality of food for grazers and higher trophic levels. | Relative importance of bottom-up vs. top-down controls on food web. Influence of habitat changes. Feasibility of management? |
| Habitat loss | Nutrients. Freshwater flows. Light, turbidity. Physical disturbance and elimination. | Can restoration of habitat facilitate recovery of key processes and native species? | Restoration uncertainties: What is manageable against a changing baseline [climate change, invasive species, declining sediment inputs]? |
| Harvest and fishing | Top-down | Implications for fisheries. | How to manage harvest for sustainable populations and to avoid top-down effects on ecosystems [sustainable production, desirable water quality, and habitat]. |
| Introduced species | Alteration of food webs and nutrient cycling. Alteration of food availability. Changes in predation. Change in physical habitat from macroflora. | Survival and management of native species. Fate of restoration actions. | Predicting success of invaders and their ecological implications? Life cycle of invasive species: can vulnerabilities be found? Controlling inputs and managing habitat for optimal production of native species. |
| Nutrients (nitrogen and phosphorus) | Nitrogen/phosphorus loads. Flows. Temperature. | Nutrient input reductions. | Determine nutrient input and flow thresholds for eutrophication and algal bloom formation and macroflora. Roles of ratios and forms of nutrients in determining community composition. |
| Passage impediments | Dams. Migration barriers. Water diversions. | Inability of species to utilize former habitats. | What species most affected by diversions? Feasibililty of management? |
| Toxic chemicals | Inputs of selenium, mercury, pesticides. | Concentrations not declining and could increase. | Selenium: San Joaquin River inputs to the Bay? Mercury: methylation increase from wetland restoration? Pesticides: How many areas of high concentration and where? Improved management. |
aDrivers listed in alphabetical order.
SOURCE: Modified from Healey et al. (2008a).
The Bay Delta Conservation Plan (BDCP) Independent Science Advisors (BDCP 2007) identified two fundamental environmental gradients that control physical characteristics of habitat for various species (Figure 3-2). While the salinity gradient has always been oriented along the axes of the major rivers flowing through the delta, elevation gradients existed at a number of spatial scales. At the largest scale, there is a decrease in elevation and slope along the river channels and banks from upstream as they enter the delta, toward the bay. At the reach scale, the high natural river levees resulted in a decrease in elevation away from the channel into floodplain (upstream) and tidal marsh (downstream), and these “cross-channel” gradients were multiplied by the complex system of river and tidal drainage channels that previously occupied the delta.
Today, the network of delta levees has substantially reduced the area exposed to the tides to about 618 square miles (Culberson et al. 2008). The drainage density within the delta has been reduced and is restricted to deep subtidal channels, resulting in a limited array of environmental gradients within the delta. Natural high land (e.g., river levees) has been essentially eliminated, as have shallow channels. Tidal and riverine flow,
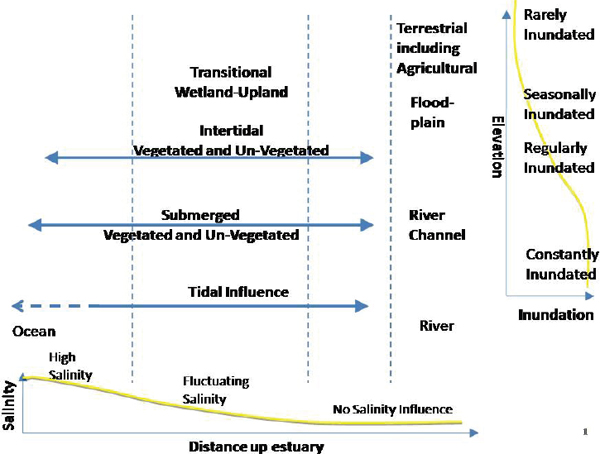
FIGURE 3-2 Horizontal and vertical gradients that control environmental conditions in the delta.
SOURCE: BDCP (2007).
across the salinity gradient, is confined to channels that do not drain at low tide. Flooded delta islands (e.g., Franks Tract, Mildred Island, and Liberty Island) are now lower than the marshes and channels in those areas would have been prior to drainage.
Isolated areas of naturally inundated wetland still exist in the delta (most of the wetlands in Suisun marsh are actually semi-impounded and their inundation regime does not therefore reflect the environmental conditions of naturally inundated wetlands). Forested floodplain with natural inundation regime is now limited to the Cosumnes River, and Rush Ranch in Suisun Bay is remnant salt marsh at the lower end of the system. Because tules (Schaoenoplectus spp.) do not require substrate drainage and can grow at elevations as low as ~0.5 m mean lower low water, tule patches exist in remnant midchannels islands and around the margins of some flooded islands. Tules have a low salt tolerance, but current water management that keeps the delta fresh for conveyance purposes allows tule wetlands to extend to the margins of Suisun Bay. Their ability to colonize into the subtidal
zone means that bare intertidal flats, which may have historically existed throughout the delta in areas periodically influenced by salinity incursion, have essentially been eliminated except in Suisun Bay. Tules can effectively dampen wave action (e.g., Augustin et al. 2009) and thus limit resuspension of sediment in shallow subtidal areas within the delta. Accordingly, the only areas where wind waves routinely resuspend sediments and provide high turbidity levels are in Suisun Bay. Ruhl and Schoelhammer (2004) found that this effect was accentuated by the storage of highly erodible sediments on mudflats in Honker Bay. If such sediments are deposited in areas colonized by tules, resuspension would be limited. Thus, the changes in elevation gradients within the delta have limited the occurrence of wetlands of various types and shallow turbid subtidal environments.
Physical Environment: Flows and Salinity
The committee’s first report, A Scientific Assessment of Alternatives for Reducing Water Management Effects on Threatened and Endangered Fishes in California’s Bay Delta (NRC 2010), dealt with aspects of flows, notably Old and Middle River (OMR) flows and X21 positioning that are specific to two biological opinions issued by the Fish and Wildlife Service (FWS) and the National Marine Fisheries Service (NMFS) to protect listed fish species, the delta smelt, and Chinook salmon. In what follows, we discuss flow effects on the aquatic resources of the bay delta more generally, aiming to set existing knowledge about these flow effects in the same framework as other stressors such as contaminants, nutrient inputs, and invasive species. To do so requires that one consider first how flow affects organisms and processes, in which cases it is anthropogenic changes to flows, volumes, timing, and paths that are the stressor(s). As discussed below, flow volumes and timing (i.e., the hydrograph) affect the temporal and spatial variability of the physical environment, a term we use to mean environmental variables like salinity, turbidity, turbulence level, as well as elements of habitat connectivity associated with horizontal transport (Cloern 2007, Cowen et al. 2006) and vertical turbulent mixing (Lucas et al. 1998). By flow paths we mean transport of organisms and materials through various regions of the bay delta, including the entrainment of listed species by the water project pumps. The issue of entrainment is dealt with below.
The distinction between these two types of flow effects on organisms, the food web, and thus on the ecosystem more generally is important in that sustainable approaches to reducing the effects of flow stressors may be quite different. In particular, the issue of flow paths appears amenable to engineering solutions: With the correct water engineering, entrainment
![]()
1 See page 20 for a definition of X2.
effects might be eliminated, allowing the maintenance of current diversion volumes, or possibly even permitting increased diversions. In a similar fashion, the problem some fish species have because of altered flow paths might be solved via strategies such as using information about when specific fish species (at various life stages) are at risk of entrainment and, with the aid of modeling, modify pump operations to reduce entrainment.
In contrast, the effects of flow on the physical-chemical environment, most notably the salinity field and its concomitant influences on circulation and transport (Monismith et al. 2002, MacCready and Geyer 2010), do not appear amenable to engineering solutions other than to use specific flow standards tied to water year type and variability, that is, standards like the X2 standard developed by the Environmental Protection Agency in 1995,2 which has subsequently been used as a basis for developing a variety of standards, including the recently proposed and litigated Fall X2 standard as well as X2 rules as described in State Water Resources Control Board (SWRCB) decision 1641.3 In this case, the development of regulations to maintain salinity gradients relies on the central hypothesis that the environmentally optimum approach is to try and mimic the shape of the natural hydrograph albeit at a lower level—in other words, to make the system slightly drier than it would be naturally, but maintain the overall pattern of flow. The key conceptual model on which this hypothesis is based is that the current ecosystem is adapted to the presence of a particular seasonal variability in flow, which certainly has varied on evolutionary time scales (Ingram et al. 1996), as discussed by Moyle et al. (2010). As a consequence, many species have life strategies that depend on particular features of flow variability, such as the transport of eggs into suitable habitat at the correct time or the aggregation of ichthyoplankton into regions of higher food availability by gravitational circulation (Arthur and Ball 1979, Kimmerer et al. 1998).
Also, the California SWRCB has recently been actively engaged in developing regulations for various aspects of flows and diversions,4 an effort that has been backed up by a detailed examination of the manifold ways in which flows affect bay-delta biota discussed in the technical report presented by Fleenor et al. (2010) to the SWRCB.
![]()
2 Federal Register, Volume 60, Number 244.
3 D1641 was finalized in March 2001.
4 Development of Flow Criteria for the Sacramento-San Joaquin Delta Ecosystem, August 3, 2010.
Hydrologic Factors
The term “flow” encompasses a broad range of effects in the bay-delta estuary. We define flow here as freshwater flow, something that has multiple components and in the context of the delta can best be thought of in terms of four major components:5 Sacramento River inflow; San Joaquin River inflow; net delta outflow, the total time averaged flow past Chipps Island at the western edge of the delta; and in-delta diversions, most notably the state and federal water projects. These four are not independent and are represented in an average sense (to a good degree of approximation):6
Net delta outflow = Sacramento River inflow
+ San Joaquin River inflow – In-delta diversions
Both of the river flows include the effects of reservoir operations (storage and releases) and diversions in and upstream of the delta, for example, the Hetch Hetchy Aqueduct, which transports Tuolumne River water to the San Francisco Bay Area. Because tidal flows at the eastern end of Suisun Bay are generally an order of magnitude larger than are mean flows (e.g., Walters et al. 1985, Monsen 2000), net delta outflow is a calculated rather than measured quantity.
One can look at anthropogenic changes in the hydrology of the bay delta by comparing measured hydrographs with the “unimpaired” hydrograph, that is, the hydrograph that would have been observed in the absence of the water projects, but including the present delta configuration. For example, in their presentation to the SWRCB, Chung and Ejeta (2011) more generally note that, as currently calculated, unimpaired flow is based on the hydrologic behavior of the system at present, rather than the system as it existed before dams, flood control levees, and so on were built. For this reason, the calculated unimpaired flow might actually be significantly different from what actually took place prior to development. Consequently, unimpaired flow should be treated as an approximate upper bound on the natural flow. To our knowledge, an appropriate lower bound has yet to be defined.
Finally, besides a reduction in the overall volume of freshwater entering the bay, the timing of flows has also been altered, with peak flows now occurring earlier in the year (February and March) than they would in the absence of water resources development. Here too, the change is not unequivocally due to water resources development: rather, it also appears
![]()
5 Besides these flows there are also the East Side streams; see http://www.water.ca.gov/dayflow/.
6 A full water balance for the delta includes groundwater–surface water exchanges as well as evapotranspiration by delta vegetation (see, e.g., Fox 1987).
that precipitation in the Central Valley watersheds is increasingly taking the form of rain rather than snow (Dettinger and Cayan 1995, Cloern et al. 2011), a pattern that also tends to shift the hydrograph peak earlier in the year. Thus, to a first approximation, the flow stressor is defined by changes in hydrology, both in volumes and timing.
Flow Effects on the Physical Environment
In conjunction with mixing from the tides, freshwater flow determines the spatial structure of the salinity field, via the relationship between flow and the position of X2. (The position of X2 is a distance scale—kilometers upstream, or east of the Golden Gate Bridge—for salinity intrusion. Thus, if X2 is at 70 km, it is 70 km east of the Golden Gate Bridge.) The reason is that at steady state the tendency for freshwater flow to carry salt out of the estuary is balanced by the tendency for gravitational circulation and tidal dispersion to carry salt upstream toward the delta. As a result of this balance, the mean position of X2 is proportional to the net delta outflow raised to the minus one-seventh power (Monismith et al. 2002), meaning that it takes much higher flows to move X2 when X2 is farther to the west, or nearer the Golden Gate Bridge, than when it is farther to the east (Figure 3-3). For example, to position X2 at 72 km (opposite Honker Bay), a flow
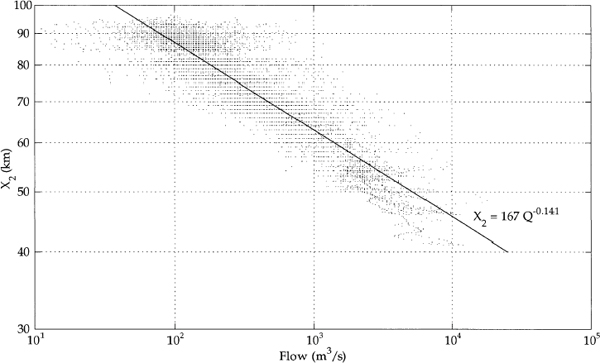
FIGURE 3-3 The position of X2 in kilometers east of the Golden Gate Bridge as a function of flow.
SOURCE: Monismith et al. (2002).
of approximately 14,000 cubic feet per second (cfs) is required, whereas to position X2 at 82 km (at the confluence of the Sacramento and San Joaquin rivers) requires 5,500 cfs. The difference in the total volume of outflow between these two positions for 1 month amounts to 500,000 acre-ft. When the position of X2 is at 72 km, it requires 1,350 cfs to move X2 1 km downstream, whereas when the position of X2 is at 82 km, it requires 470 cfs to do so. As context, the tidal excursion in Suisun Bay and the western delta is of the order of 10 km.
The location of X2 affects several key aspects of the physical environment. First, as reported by Jassby et al. (1995), the local depth-averaged salinity at any distance, x, from the Golden Gate can be estimated approximately as the product of the salinity at the Golden Gate and a function of the ratio of x to the position of X2 (Figure 3-4). Thus, a given value of the distance of X2 from the bridge will determine the salinities in a wide variety of habitats (i.e., the channels of the estuarine portions of the Sacramento
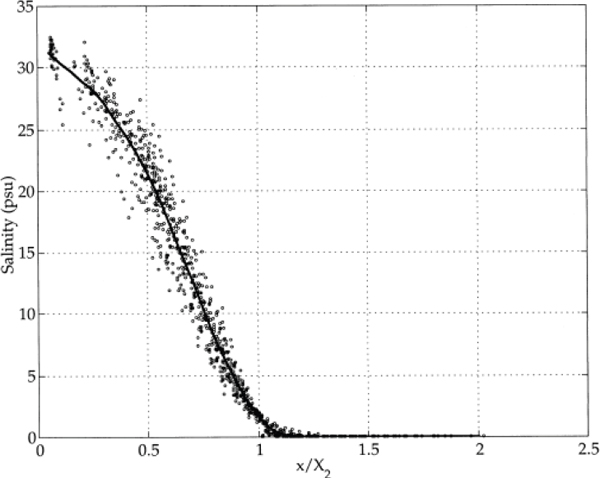
FIGURE 3-4 Depth-averaged salinity as a function of the ratio x/X2.
SOURCE: Monismith et al. (2002).
and San Joaquin rivers, or the shallows of Grizzly and Honker Bay). This relationship was considered by Kimmerer et al. (2009), who used a 3D circulation model to look at the volume of habitat at a given salinity and depth that might be expected on statistical bases to be used by various species at different life stages.
Second, the position of X2 also specifies the strength of the salinity gradient and the strength of gravitational circulation and the intensity and persistence of vertical density stratification (Monismith et al. 2002). Upstream of X2, gravitational circulation is absent, whereas downstream of X2 it varies both tidally and subtidally (Stacey et al. 2001). As suggested by a number of studies (e.g., Laprise and Dodson 1994, North and Houde 2001), gravitational circulation can play an important role in the retention of estuarine species that otherwise might be swept out of the estuary by tidally averaged flows (e.g., Orsi 1986). Indeed as described by Arthur and Ball (1979), because it is the boundary between the presence and absence of gravitational circulation, the low-salinity zone (LSZ) near X2 may be a region of concentration of zooplankton as well as larval fish. However, this conceptual model may be applicable only to conditions existing when X2 is sufficiently far upstream to keep the LSZ in the more channelized sections of the Sacramento and San Joaquin rivers. When the LSZ is in Suisun Bay, energetic horizontal mixing associated with tidal motions over the highly variable bathymetry (Ridderinkhof and Zimmerman 1992, Burau et al. 1993) may weaken the tendency to accumulate organisms in the LSZ. This remains to be evaluated with 3D modeling. Moreover, particle tracking calculations of model zooplankton in the St. Lawrence estuary reported by Simons et al. (2007) show that swimming behavior can substantially increase retention of organisms in an energetic LSZ (see also Bennett et al. 2002).
The vertical structure of the water column is also related to the position of X2. Upstream of X2, the water column is generally unstratified, whereas downstream it stratifies and destratifies tidally, with stratification lasting through the tidal cycle when X2 is sufficiently far downstream and/or at neap tides (Stacey et al. 2001). Moreover, the strength and persistence of stratification varies inversely with the position of X2, such that the estuary is less stratified when X2 is in the delta than when it is in Suisun Bay (Monismith et al. 2002). This is important because stratification strongly weakens vertical, turbulent mixing, potentially decoupling the benthos, and benthic grazing (Cloern 1982) from the rest of the water column, and enabling motile phytoplankton to remain in the near-surface photic zone (Koseff et al. 1993) rather than being mixed into the deeper parts of the water column where respiratory losses of biomass can be larger than gains from photosynthesis. In this way, the physical environment of the bay delta is fundamentally affected by flow in ways that may significantly affect primary production and food-web dynamics in regions downstream
of X2 (whether or not X2 is found in the delta or downstream in Suisun or San Pablo bays).
Finally, the position of X2 may be important to the likelihood of entrainment of organisms in that when X2 is upstream of the confluence of the two rivers, organisms that associate with the LSZ (e.g., larval fish), are more likely to be within the region of influence of the pumps (Kimmerer 2004). However, the positioning of X2 far upstream may also occur with large exports as well as small outflows.
Flow Effects on Aquatic Resources: Primary Production in the Delta
Net delta outflow is thought to influence the residence time of materials in various regions of the delta (Monsen 2000, Monsen et al. 2007), and so should influence primary production in the delta (Jassby and Powell 1994, Jassby 2008). The concept of residence time in the delta is complicated by two factors. With the exception of Mildred and Liberty islands and Franks Tract, water is not well mixed on the scale of the delta and so no single residence time can be defined, and mixing by the tides is energetic, especially on the Sacramento side of the delta (Monsen 2000, Monismith et al. 2009) so that, even without any freshwater flow, there would be exchange between the delta and San Francisco Bay. The only examination of the effect of inflow on residence time that we are aware of is that by Kimmerer and Nobriga (2008). Using the particle tracking capability of DSM2, a 1D network model, they found that, in the northern delta, computed residence times matched or were shorter than the overall hydraulic replacement time of the delta (the delta volume divided by total inflow). Pointing to the complex nature of transport processes in the delta, computed residence times in the central and southern delta were larger than the hydraulic replacement time, did not vary monotonically with flow, and were affected by exports as well as inflow.
The connection between physical transport and primary production was examined by Jassby et al. (2002), who found that, as expected, increased inflow decreased phytoplankton biomass in the delta (as measured by chlorophyll a). Note that inflow is the correct flow metric since all the water that enters the delta must leave, mostly via outflow to the bay or by export from the pumps. Jassby (2008) extended these results showing a dramatic shift downward in the biomass-flow relation between 1980 and 2000. Besides flow, geometry of the delta can also influence residence time. For example, Monsen et al. (2007) found that placement of the Head of Old River Barrier, a temporary barrier designed to reduce entrainment of outmigrating salmon smolts in the San Joaquin system, significantly reduced residence time in the San Joaquin ship channel.
A more subtle effect of transport on primary production is that trans-
port can couple regions of high productivity with regions that are strong sinks for primary production due to benthic grazing (Lucas et al. 2002), such that increasing residence time can reduce the accumulation of phytoplankton biomass. As an aside, this points to a possible problem with proposals (e.g., in the BDCP) to increase primary production in the system by increasing shallow water habitat: if that shallow water habitat includes a significant biomass of benthic grazers, it may become a net sink for primary production and so will decrease the total phytoplankton biomass available for pelagic grazers like zooplankton. Finally, mixing and transport may not act equally on all types of phytoplankton. In particular, grazing may have a much smaller effect on positively buoyant cyanobacterial genera like Microcystis than it does on negatively buoyant species such as the various diatoms that are thought to be good food for zooplankton.
Flow Effects on Aquatic Resources: Effects of the Position of X2
Evidence demonstrating the effects of flow on bay and delta biota was presented by Jassby et al. (1995), who used Interagency Ecology Program (IEP)7 data from the period 1968-1991 to show that the abundance (biomass) of a number of organisms, including the total production of particulate organic carbon by phytoplankton in Suisun Bay, the shrimps Neomysis mercedis and Crangon franciscorum, and several fishes, for example, starry flounder (Platichthys stellatus), striped bass (Morone saxatilis), and longfin smelt (Spirinchus thaleicthys), but notably, not delta smelt nor the key zooplankton Eurytemora affinis, was dependent on the values of the position of X2 averaged over various parts of the year. The averaging periods, which ranged from 4 months to a year (see Table 1 in Jassby et al. 1995), were chosen by considering when flow variations might have an important impact, given known life histories of each organism.
The results of Jassby et al. (1995) do not exclude the importance of entrainment. Indeed they showed that for the particular case of striped bass, a better prediction of population size could be had by including diversions as well as the position of X2 in the statistical model used to represent spring striped bass survival. Interestingly, the more complicated model had larger uncertainty in terms of determining the position of X2 that would be required to the median observed level of survival.
Kimmerer et al. (2009) reexamined the results of Jassby et al. (1995), considering separately the period before 1987 and the period from 1987 to 2006, with 1987 chosen as the approximate start of the Corbula invasion of San Francisco Bay. For most species considered (but not delta smelt), Kimmerer et al. (2009) found that the slopes of the abundance–X2-position
![]()
relationships were similar for the two periods, although absolute abundances for a given value of the position of X2 in some cases (e.g., longfin smelt) were reduced. Using modeled salinity fields and observed distributions of each species, Kimmerer et al. (2009) defined habitat indices for each species that also varied with the position of X2 (Figure 3-5). Using these indices, they found that only for American shad (Alosa sapidissima) and striped bass were the abundance–X2-position and habitat–X2-position relations consistent, leading them to conclude that only for these species was habitat the means by which X2 position influenced abundance. For the other species for which a connection to X2 was inferred, the mechanisms behind observed X2-position–abundance relations remained to be determined.
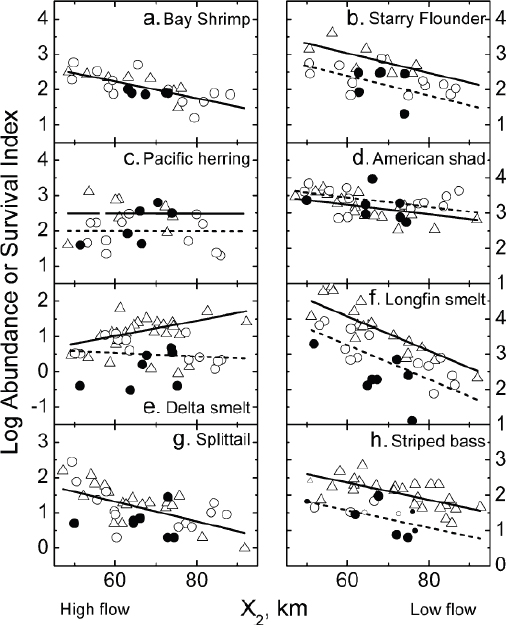
FIGURE 3-5 X2-abundance relationships for fish and shrimp. Symbols indicate data up to 1987 (triangles and solid lines); 1988-2006 (open circles and dotted lines); and 2000-2007 (solid circles). The panels for striped bass and Pacific herring plot survival indices; all the others show log abundances.
SOURCE: Kimmerer et al. (2009).
For example, for starry flounder and bay shrimp, organisms that “recruit from the coastal ocean along the bottom into the estuary,” Kimmerer et al. (2009) suggested that, as the distance upstream of X2 decreases, gravitational circulation strengthens, and the transit time of organisms from offshore hatching sites to their estuarine rearing grounds would be reduced, presumably increasing survival. For the Sacramento splittail (Pogonicthys macrolepidotus), increased abundance when X2 is downstream may be from an increase in floodplain spawning habitat because high-flow years, when X2 is farther downstream, tend to involve flooding of the Yolo Bypass (Feyrer et al. 2006). Most recently, in the context of the pelagic organism decline (POD; see further discussion below), Mac Nally et al. (2010) reanalyzed the IEP data for 1967-2006, finding that
1. the position of X2 in the spring (“spring X2”) strongly influences the abundance of mysids, longfin smelt, and calanoid copepods; fall X2 (referring to the position of X2 in the fall, by analogy with spring X2) strongly affects striped bass; and winter X2 has a weaker effect on delta smelt; and
2. spring and fall X2 did not appear to have any statistical relation to delta smelt abundance, although it was related to smelt distribution.
Thus, while the mechanisms behind the influence of position of X2 on the abundance of a variety of biota remain hypothetical, the statistical relations reported in several papers show that abundance of a number of species at different trophic levels found in the delta and San Francisco Bay is higher when X2 is farther downstream. This implies that sufficient reductions in outflow due to diversions would tend to reduce the abundance of these organisms.
Physical Environment: Turbidity
Sediment particles, phytoplankton, and other suspended materials in the water column causing turbidity affect light penetration in the waters of the bay delta, as do colored dissolved materials. The degree of light penetration limits primary production by phytoplankton (e.g., Cole and Cloern 1984) and submerged aquatic vegetation (SAV) (e.g., Carr et al. 1997) and may shape a wide range of fish behaviors such as feeding (Baskerville-Bridges et al. 2002), since small fish or fish larvae are at risk to predation by visual predators.
On tidal time scales, sediment concentrations in the bay and delta generally reflect a local balance between erosion, settling, vertical turbulent mixing, and horizontal advection (Krone 1979; McDonald and Cheng 1997). In the deeper channels, turbulent processes (i.e., erosion and verti-
cal mixing) are largely due to tides, whereas in the shallow shoals, wind-driven waves are dominant (May et al. 2003). In many estuaries a turbidity maximum (estuarine turbidity maximum or ETM) forms in the LSZ where near-bottom upstream transport by gravitational circulation leads to particle retention (e.g., Geyer 1993), a mechanism thought for some time to be important in the bay delta (Arthur and Ball 1979), although more recently it has become clear that there are multiple ETMs in Suisun Bay, all tied to local bathymetric features rather than to the structure of the salinity field (Schoellhamer 2000). This distinction is important because much of the initial basis for an X2-flow standard was based on positioning of such an ETM in Suisun Bay (Williams and Hollibaugh 1987).
Given the high turbidity of much of the bay and delta (the Secchi disk depth—a measure of visibility—is typically less than 1 m), planktonic primary production probably is light limited (Cole and Cloern 1984) such that relatively high levels of nutrients have not resulted in algal blooms. Indeed, the delta is one of the least productive temperate estuarine ecosystems (Jassby et al. 2002). Consequently, physical processes such wind (May et al. 2003), which affect suspended sediment concentration, can limit the formation of algal blooms. Importantly, the dominant paradigm of light limitation, which has been well supported by extensive observation, has meant that until recently (Dugdale et al. 2007) little attention has been given to the role of nutrients in primary production. Primary production is discussed in detail below.
Analyzing 36 years’ of data, Feyrer et al. (2007) inferred that delta smelt, and age-0 striped bass, were more likely to be found in turbid water, although turbidity explained only 13 percent of the variance in delta smelt occurrence (not abundance). However, Mac Nally et al. (2010) found that delta smelt abundance was not related to turbidity, although the effect of turbidity on occurrence and abundance may be different if the amount of appropriately turbid (and saline) habitat does not limit current delta smelt populations. The effect of turbidity on delta smelt populations may take other forms. Grimaldo et al. (2009b) argue that the appearance of significant numbers of delta smelt at the export pumps is related to the appearance of the first flush of turbid water through the delta and that, accordingly, monitoring of turbidity could be used as a basis for guiding pump operations. On the other hand, the evidence for this behavior presented in their paper (e.g., Figure 6) appears rather weak in contrast to observed relationships between delta smelt salvage and zooplankton abundance or negative OMR flows (Grimaldo et al. 2009b, Figures 7 and 8).
To cast turbidity as a stressor, it is necessary to examine changes and trends in turbidity, namely the fact that turbidity is decreasing with
time (Schoellhamer 2011; Wright and Schoellhamer 2004). This has been hypothesized to be a result of decreased sediment supply to the estuary because a significant fraction of sediments that would enter the system naturally are now trapped in upstream reservoirs (Schoellhamer 2011) and in stabilized floodplains. This hypothesis is reinforced by the fact that changes in bathymetry over the past 100+ years (Jaffe et al. 1998) indicate that the system is now net erosive, indicating that the sediment supply into the estuary is exceeded by net exports to the ocean.
These changes are perceived as a stress to pelagic organisms, although the importance of current practices and water project operations (aside from storage of sediments) is less clear because much of the suspended sediment that determines turbidity today was originally deposited in the system in the 19th century through now-banned mining practices (Schoellhamer 2011).
In summary, like salinity, turbidity is a fundamental aspect of the physical environment of the delta, and so systematic, long-term changes in turbidity appear to be important to the ecosystem. Increased clarity should result in increased primary production by both phytoplankton and by SAV. For phytoplankton, this might mean a shift away from dominance by light limitation, toward more nutrient-limited conditions. Given that the connection between turbidity and primary production by phytoplankton is well known, this response of the system should be straightforward to predict. However, in situ it is probably a highly variable trade-off between periods of higher and lower turbidity, leading to highly variable light versus nutrient-limited conditions.
Increasing clarity might also favor negatively buoyant species like diatoms over positively buoyant cyanobacteria like Microcystis that can do well in turbid environments, and so it could act to reduce the production of cyanobacteria. For SAV, it appears that increasing clarity may yield increases in nonnative species, most notably Egeria. This trend may play an important role when attempting habitat restoration.
Increasing clarity will also act to decrease the amount of suitable habitat for small fish that favor shallow, turbid waters, and make those fish (albeit to an unknown extent) more vulnerable to predation. This effect may be more pronounced in the deeper delta channels rather than in the wave-mixed shallows of Suisun Bay. However, while there are compelling biological reasons to conclude that turbidity is important to fish, the statistical evidence connecting turbidity to abundance is somewhat weaker than that connecting flow to abundance. Thus, at present there is insufficient evidence to conclude that turbidity can be used or manipulated to lessen impacts of diversions on fish.
Physical Environment: Temperature
Water temperatures at any point and time in the delta are determined by heat exchanges with the atmosphere, by long- and shortwave radiation, by horizontal advection by currents (tidal and nontidal), and by vertical, turbulent mixing (Fischer et al. 1979, Wagner et al. 2011. The overall setting of the bay delta involves cold temperatures at the ocean end (particularly during upwelling) and at the riverine end, with warmer temperatures in between. Like salinity, temperature is affected by flow (i.e., net delta outflow for the bay, and inflows and net delta outflow for the delta). For example, modeling and observations reported by Monismith et al. (2009) show that, for the San Joaquin system, net flow through the system acts to push the region of maximum temperature downstream toward the ocean. Regulation of temperature primarily is focused on river sections downstream of dams, where selective withdrawal of cold water can be used to help keep instream temperatures sufficiently cold for salmonids. Further discussion of temperature is in Chapter 4.
Physical Environment Conclusions:
The Management Dilemma of Habitat Versus “Plumbing”
The structure of the bay-delta ecosystem is related to the structure and variability of its physical environment. This physical environment has been significantly altered by the development of California’s water resources, most notably by changes in flows into and through the system. However, in practical terms, some elements of these alterations are more amenable than others to actions aimed at improving ecological rehabilitation of the ecosystem. For example, other than in riverine regions close to dams, we have little ability to affect temperatures, except through flow. Aside from the fundamental issue of storage of sediments in reservoirs, turbidity (and its variation) is primarily a result of natural forces. Importantly, given that diversions in very wet years constitute a small fraction of the unimpaired flow in winter and spring, it appears that important aspects of variability of flow are outside the control of water project operations. Nonetheless, human use of water does have significant influence on freshwater flow much of the time. Thus, some form of flow management is of paramount importance for ecological rehabilitation.
In considering flow management, it is critical to recognize that the issues raised by the relationship between the position of X2 and abundance of many species are fundamentally different than those associated with entrainment of fish. In principle, entrainment of fish is a problem localized to the delta that can (optimistically) be solved by changing the water engineering of the delta. In effect, the fish salvage facilities at the State Water
Project (SWP) and Central Valley Project (CVP) pumps represent the first attempts at eliminating direct entrainment effects; indeed, if the facilities were perfect and predation near the pumping facilities were negligible, 100 percent of the fish that find their way to the pumps would be saved. In reality, salvage is quite inefficient, and only a small fraction of the entrained fish survive salvage (Brown et al. 2009). A second example is the gate on the Delta Cross Channel (DCC), which can be closed to improve survival of salmon smolts on the Sacramento River side. However, closure of the DCC tends to increase salinities in the western delta, affecting water quality at the Contra Costa Water District water intake (Monsen et al. 2007). A similar alteration to the plumbing is the Head of Old River Barrier (see above). It too may have collateral negative effects by possibly increasing the entrainment of delta smelt resident in the south delta (Kimmerer and Nobriga 2008). The most radical effort of this type was a proposal by the Metropolitan Water District to build and operate two sets of gates on Old and Middle rivers, with operations tied to turbidity variations that may affect delta smelt.8
The largest alteration to the flow-path engineering, one originally contemplated in the planning of the SWP, is one designed to avoid entrainment directly by separating the diversion of Sacramento River water from the rest of the delta. If used by itself, and if screening on the intake is successful, such a facility might reduce entrainment as well as reverse mean flows in the delta that might affect fish migration.
However, the utility of these plumbing measures depends on two factors: the importance of entrainment to fish populations (see stressor section below) and the degree to which outflow from the delta into San Francisco Bay itself does not influence species abundance or other ecosystem attributes. The advantage of changes to the flow paths is that these active, engineering measures might support human use of freshwater entering the delta, while also providing some degree of environmental protection.
In contrast, flow effects that affect San Francisco Bay downstream of the delta, as might be represented in the relations between the position of X2 and abundance, are not amenable to direct engineering intervention in that the only things that can be controlled are timing and volume of flow out of the delta. Given that the position of X2 for different periods of time appears to be important for different species, one can argue that water operations should be designed to preserve as much of both the volume of outflow and the timing of that volume that would be observed in the absence of diversions (Moyle et al. 2010, SWRCB 2010). In light of the nature of the connection between flow and the position of X2, this may necessitate limiting available water supply, especially in dry years.
![]()
Nutrient Enrichment
Macronutrients (nitrogen, phosphorus, silicon) and micronutrients (trace metals and iron) are essential for supporting and sustaining primary and secondary production in aquatic ecosystems, including the delta. Microalgae, specifically phytoplankton, are dominant primary producers in the delta and lower bay systems; hence, this section focuses on them. High nutrient inputs can lead to altered community structure and proliferation of phytoplankton that may have undesirable effects on biogeochemical cycling, food-web dynamics, habitat conditions, and human health. There are numerous examples of the negative effects of nutrient overenrichment, or “too much of a good thing” (D’Elia 1987) worldwide (Schindler 1971; Smetacek et al. 1991; Vollenweider et al. 1992; Nixon 1995; Paerl 1997, 2008; Boesch et al. 2001; Cloern 2001; Elmgren and Larsson 2001; Conley et al. 2009) and in the delta (Dugdale et al. 2007, Lehman et al. 2008, Meyer et al. 2009). These include (1) increased primary production, (2) selective stimulation of harmful (i.e., toxic) algal bloom species, and (3) shifts in phytoplankton community structure to more opportunistic species that (4) induce changes in food-web structure and trophic transfer and (5) enhance the potential for bottom-water hypoxia and anoxia due to increased sedimentation of autochthonous (indigenous) organic matter.
High inputs of both nitrogen (N) and phosphorus (P) can accelerate estuarine eutrophication (Nixon 1995, Boesch et al. 2001, Elmgren and Larsson 2001, Conley et al. 2009, Paerl 2009), with P playing a more important role in the freshwater regions and N playing a more dominant role in marine systems (Nixon 1995, Paerl 2009). However, in transitional environments like estuaries, both N and P play interactive controlling roles (Fisher et al. 1992, Paerl 2009). Both the amounts and ratios of N and P inputs and resultant concentrations can determine the structure and functioning of primary producers. The various chemical forms of these nutrients can play additional roles in modulating community responses. Finally, there are synergistic and antagonistic interactions among limiting nutrients. For example, N and P co-enrichment often leads to greater degrees of bio-stimulation than N or P alone (i.e., they may be co-limiting) and the effects of N enrichment may be amplified by parallel iron (Fe) enrichment, since N assimilatory enzymes require Fe as a structural component, and energy yielding biosynthetic pathways requires Fe as a cofactor.
Alpine and Cloern (1992), Cloern and Dufford (2005), and Jassby (2008) pointed out that in turbid, highly tidally mixed, well-flushed, nutrient-enriched estuaries like San Francsico Bay and the delta, light availability, flushing rates (i.e, water residence time), and filter feeding assume important, and at times dominant, roles in limiting phytoplankton production. In well-flushed regions of the bay and delta, both N and P are often
plentiful (i.e., exceeding the half-saturation constants9 for growth), and N:P supply ratios or different chemical forms have little effect on shaping phytoplankton community structure and function. However, in some regions of the bay delta where tides are weaker (e.g., the southernmost reach of South San Francisco Bay or the southern interior delta) water residence time may be long enough (especially during low-flow periods) for nutrients to be thoroughly assimilated (Cloern 2001), leading to biomass increases (unless grazing exerts a strong control). Under these conditions, nutrient limitation is most likely to occur, and nutrient enrichment could impact the species composition and functioning of primary producers and consumers. Given that water residence times can vary on short- (diel) and longer-term (seasonal, interannual) time scales, nutrient limitation might be intermittent rather than continual. This possibility should be investigated for the delta.
Additionally, in many parts of the delta, where flows are weaker and water withdrawals and diversions have taken place, flow and residence time have been altered (Lucas 2009). Recent studies in these habitats (Frank’s Tract, Mildred Island) have shown that such hydrologic alterations can affect phytoplankton community structure (Lucas 2009, Lucas et al. 2009). In addition, delta geomorphology, and human changes therein, can affect flow, residence time, and potentially nutrient assimilation, primary production, and phytoplankton growth and composition. Monsen et al. (2007) provided an example of how the placement of a barrier in the south delta radically changed flushing times and water quality.
When flow and flushing are reduced and water residence increases, phytoplankton will have more time to assimilate nutrients and build up biomass (as blooms) before being transported out of the system. This scenario benefits phytoplankton in general, and more specifically those species that have generally slow growth rates, since under the influence of reduced flushing (longer residence time) these species will more effectively compete with faster-growing species for nutrients and other resources. Most cyanobacteria (blue-green algae), including harmful bloom–forming types, exhibit relatively slow growth rates (Paerl and Huisman 2009, Paerl et al. 2011). Hence, reduced flow and flushing conditions tend to favor cyanobacteria, especially if nutrient supplies are adequate to sustain blooms (Paerl 2008, Paerl and Huisman 2009). In recent years, the non-N2 fixing, potentially toxic bloom–forming cyanobacterium Microcystis spp. has increased in dominance in slow-moving fresh to oligohaline waters of the upper delta (Lehman et al. 2008). This genus appears to have benefitted from the combined effect of reduced flushing (increased residence time), possibly warmer
![]()
9 In algal physiology, the half-saturation constant is used to describe the general affinity of an enzyme for a substrate or nutrient, which allows one to estimate whether an organism is operating under nutrient-limiting or nutrient-saturating conditions.
water conditions (which would enhance growth rates), more intense stratification, and increases in nutrient loading. Microcystis is indicative of nutrient-enriched conditions worldwide (Reynolds 1987, Paerl 2008). In particular, this genus tends to dominate in waters that are receiving excess N, since it is a non-N2 fixer and hence relies on externally supplied forms of N. Their recent increase appears to be due to the combined effect of increased residence time and excessive N loading (possibly combined with a warming trend), which is conducive to Microcystis bloom formation. In addition, colony-forming cyanobacterial bloom genera like Microcystis are not readily grazed by crustacean zooplankton or benthic infauna because they cannot be effectively filtered, and they produce toxic compounds that can deter grazers and they form surface scums, which cannot be accessed by benthic and subsurface planktonic filter feeders (copepods, cladocerans, invertebrates, and fish larvae).
There are geographically diverse examples that point to excessive N inputs as a factor promoting Microcystis blooms (Paerl et al. 2011). While excessive N inputs may help stimulate bloom formation, P supplies must also be available. Therefore, while there is evidence for N overenrichment, P inputs should also be examined as a possible secondary nutrient stressor that affects ecosystem structure and functioning.
Cloern (presentation to the NRC committee, July 2010) pointed out that excessive N loading may also be problematic in South San Francisco Bay, which can have dry weather residence times of several weeks (Gross et al. 1999) and is prone to harmful (i.e., potentially toxic) dinoflagellate blooms, which have recently appeared in this part of the bay.
With respect to the influence of different chemical forms of nutrients as possible stressors on the delta system, it has been proposed that the reduced form of N, ammonium, may play a selective role by inhibiting nitrate utilization and growth of diatoms in mesohaline to full-salinity regions of the delta and downstream bay regions (Dugdale et al. 2007). This scenario would depend on whether N is even limiting in this region, which has been questioned by Cloern and colleagues (e.g., as opposed to light availability, flushing and transport, and grazing as potential factors controlling phytoplankton growth) (Cloern 2001, Cloern and Dufford 2005). The ammonium inhibition argument is based on mostly oceanic observations of a strong preference for nitrate as the N source in diatom populations, and on laboratory observations that relatively high levels of ammonium (>4 μM) can inhibit the uptake of nitrate in diatoms. These observations have led Dugdale and colleagues (2007) to propose that ammonium discharge from upstream wastewater treatment plants (specifically the Sacramento Waste Water Treatment Plant or SWWTP) may be high enough to cause inhibition of nitrate uptake by diatoms in downstream waters (e.g., Suisun Bay) (Dugdale et al. 2007). If common and widespread, this type of inhibition
affects the food web and nutrient and carbon cycling, since diatoms are considered a good food source for most zooplankton, planktivorous fish, and shellfish species. In this regard, there has been a general decline in diatom biomass since the mid-1990s (Dugdale et al. 2007), and the amount of ammonium discharged by the SWWTP (and possibly other wastewater treatment plants) has shown a parallel increase. This too would give a flow effect since increased delta inflow would tend to dilute SWWTP discharges into the Sacramento River and lower ammonium concentrations in Suisun Bay.
However, Jassby (2008), Jassby et al. (2002), Thompson et al. (2008), and Cloern et al. (2010) all pointed out that the decline in diatom biomass in Suisun Bay and other locations took place shortly after the introduction of the Asian clam Corbula, a voracious grazer capable of removing vast amounts of phytoplankton biomass. Therefore, several environmental factors correlate with the decrease in diatom biomass starting in the mid-1990s. Also, in addition to decreases in diatom biomass, other phytoplankton taxa decreased in biomass at this time, at similar locations. This latter observation would tend to support the argument that “top-down” grazing exerted by invasive benthic bivalve grazers is a major control of phytoplankton biomass at these locations. Finally, it is exceedingly difficult to attribute specific ammonium supplies and concentrations in the lower delta and San Francisco Bay to the SWWTP, which is more than 100 km upstream from these locations. It is likely that ammonium, as well as other bioreactive N compounds released from the plant, go through numerous biogeochemical transformations during their travel time in the river and upper bay delta. Therefore, total biologically available N (ammonium, nitrate/nitrite, dissolved organic N) discharged from SWWTP and other anthropogenic sources should be included when considering N input reductions aimed at stemming unwanted symptoms of eutrophication (e.g., cyanobacterial blooms in the upper delta and other nutrient-sensitive regions of the San Francisco Bay, e.g., South San Francisco Bay). The role of ammonium in favoring an invasive species and thus structuring the pelagic community (Glibert 2010, Glibert et al. 2011) is discussed further below with other effects of nonnative species.
Conclusions
When physical conditions permit (i.e., increased residence time, adequate clarity, elevated temperatures, and enhanced vertical stratification), nutrients can play a role in the control of phytoplankton production and in bloom formation and persistence in parts of the delta system. Nitrogen appears to be the nutrient most likely to influence bloom formation, although a potential secondary role of P should not be ignored. Therefore, there is
agreement that N input reductions will help ensure optimal water quality conditions in the delta and possibly parts of San Francisco Bay (South Bay). There appears to be less certainty as to whether reducing one form of biologically available N is preferred over another (e.g., nitrate vs. ammonium vs. dissolved organic N). Because different forms of N are biologically available and readily cycled between the water column and sediments, the prudent approach is to reduce the impacts of all forms of organic and inorganic N, which will ensure that undesirable algal bloom formation in regions prone to such events is minimized. There is less certainty as to the role P inputs play in the control of algal production and bloom formation. If P plays a role as a limiting nutrient, it is likely to be during freshwater blooms, but this has not been established. The degree to which N reductions should be practiced is at present uncertain and requires field and laboratory research (i.e., establishing nutrient-bloom thresholds using bioassays, stoichiometric analyses, N transport, and fate and cycling studies) and modeling that takes both physical and chemical forcing features, as well as the interactive effects of grazing, into consideration.
Food Quality and Quantity: Linking Environmental Stressors to Changes at Base of the Food Web
The drastic alteration of the Sacramento–San Joaquin Delta and San Francisco Bay since at least the mid-1800s has led to multiple and interacting physical, chemical, and biological changes (Healy et al. 2008b). Among the most potentially problematic changes (from biogeochemical cycling and trophic perspectives) are those at the base of the food web, namely significant changes in the structure and functioning of phytoplankton communities, the key food source supporting higher trophic levels (Cloern 1982, Cole et al. 1992, Jassby 2008). These changes have cascaded up the food web (Healy 2008, Kimmerer et al. 2008a). Filter feeders, grazers (zooplankton and invertebrate larvae), and planktonic herbivorous fish species appear to be particularly sensitive to changes in food quantity and quality. For example, growth of delta zooplankton is limited at chlorophyll a levels of < 10 μg/L (Mueller-Solger et al. 2002). There is also evidence of similar thresholds for clams. This, combined with the data of Jassby (2008) showing median chlorophyll a concentrations in the delta much less than 10 μg/L and overall declining over time, suggests that primary consumers are, at times, food limited.
Changes in the food web were among a suite of factors examined as possible causes of the recent declines in four fish species, collectively termed the “pelagic organism decline” or “POD.” The four POD species were delta smelt, longfin smelt, threadfin shad (Dorosoma petenense), and juvenile striped bass. There were several related drivers or causative factors
that caused changes in the food web that were considered. These include a decline in diatoms (preferred food source for grazers) in the Susin Bay and other areas (Dugdale et al. 2007, Cloern et al. 2010), increasing prevalence of potentially toxic and cyanobacterial bloom species, which also are of low nutritional value, in the delta region (Lehman et al. 2005, 2008, 2010), trophodynamic changes (phytoplankton and zooplankton) in the delta and bay caused by the proliferation of exotic species, most notably the overbite clam (Corbula amurensis) and Asian clam (Corbicula fluminea), and the expansion of invasive aquatic macrophytes (e.g., Brazilian waterweed, Egeria densa) affecting phytoplankton in some regions of the delta. The decline in diatoms has been attributed to excessive ammonium (Dugdale et al. 2007) and Corbula grazing (Alpine and Cloern 1992, Jassby et al. 2002, Greene et al. 2011).
Blooms of the colonial cyanobacteria Microcystis are problematic from a food-web perspective, because even though these blooms can produce large amounts of biomass, they are either avoided or not captured and assimilated by key crustacean (copepods, cladocerans) zooplankton species and invertebrate larvae that serve as a food source for numerous ecologically and recreationally important fish species (Paerl et al. 2001, Lehman et al. 2008, 2010).
Corbula amurensis, an aggressive invader, has populated the benthic regions of San Francisco Bay and the western delta to the extent where its density has reached 10,000 per m2. Its grazing capabilities are such that it is capable of quantitatively “grazing down” phytoplankton populations (Cole et al. 1992, Thompson and Nichols 1996), which appears to have led to a state change in segments of the northern bay where phytoplankton biomass exhibited a precipitous and sustained decline coincident with the proliferation of these bivalves (Alpine and Cloern 1992, Jassby 2008; J. Cloern, presentation to the NRC, July, 2010). Finally, the expansion of invasive aquatic macrophytes (e.g., Brazilian waterweed, Egeria) may also play a role in the declining dominance of phytoplankton in some regions of the delta.
The same drivers that affected the phytoplankton also affected the zooplankton. In some cases, these drivers (e.g., introduced species) directly affected the zooplankton. In other cases, the driver effects were indirect, through their direct effects on the phytoplankton that support the zooplankton. The major changes in the zooplankton are described by Kimmerer in Appendix E. Nutrients, optical properties, residence time, and invasive species also affect aquatic macrophytes, such as Brazilian waterweed; these macrophytes have large ecosystem effects, as described in the next section. Changes in nutrients affect their growth (e.g., Feijoo et al. 1996, 2002).
When taken all together, the changes in the food base from top-down grazing and macrophyte competition (above) can be viewed as alternative
hypotheses to the “ammonium inhibition” and more general N overenrichment hypotheses (see nutrient-enrichment section). Most likely, there are interacting environmental drivers at play in controlling the qualitative and quantitative makeup of food supplies at the base of the delta and bay food webs.
Conclusions
There is a need to distinguish changes in physical drivers such as freshwater discharge, turbidity, temperature, and vertical mixing as well as circulation, from chemical factors, such as nutrient enrichment and changes in nutrient supply ratios, and biological factors, including top-down grazing, as causative agents for changes at the base of the food web and the POD. All of these factors affect rates of primary production, standing stock, and composition of primary producers along the freshwater-to-marine continuum representing the bay-delta system. These diverse but often interacting drivers have been illustrated in the conceptual diagram presented by Meyer et al. (2009) in their evaluation of the role of ammonia/ammonium in food-web and biogeochemical dynamics of this complex system (Figure 3-6).
Drivers of quantity and quality of primary production of the bay-delta ecosystem include climate, hydrology “(including upstream water withdrawals and other flow modifications), human activity, loadings and types of nutrients (mainly N and P, from anthropogenic and natural sources), loadings and types of contaminants (including NH3/NH4+, NO2–, metals, pesticides and algal toxins), sediment loadings, light, and food web processes (including trophic interactions, with special emphasis on invasive species)” (Meyer et al. 2009). Because they co-occur in space and time, these drivers are highly interactive, synergistically and antagonistically, and hence should be portrayed this way. These interactions are conceptualized in Figure 3-6. Meyer et al. (2009) aptly summed up the interactive nature of these environmental controls on food source and type as follows:
These factors are interrelated in a complex web of physical, chemical and biological processes…. Climate and hydrologic variability are closely related factors that, in conjunction with human activity, influence and to varying degrees control many of the other drivers (e.g., delivery of nutrients and contaminants, changes in residence time). Therefore, climate/hydrologic variability and human activity are placed on the left in [Figure 3-6], with consequences of those factors cascading from left to right through all the other drivers and ecosystem components. The endpoints of major concern in this framework are changes in the Bay-Delta food web and populations of the POD organisms, as shown on the right in [Figure 3-6].
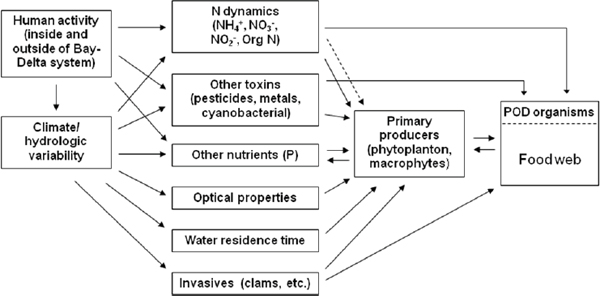
FIGURE 3-6 Conceptual framework of major drivers of water quality and ecosystem structure and function, and their relationships to the food web and POD organisms, in the Sacramento-San Joaquin Delta and Suisun Bay ecosystem. The solid arrow from the N dynamics box to the primary producers box indicates traditional processes associated with nutrient supply and uptake; the dashed arrow indicates the proposed inhibitory/competitive effect of NH4+ on uptake of NO3– by diatoms.
SOURCE: Meyer et al. (2009).
Introduced Species
The bay delta has been referred to as “the most invaded estuary in the world” (Cohen and Carlton 1995). This statement focuses on rates of invasions, i.e., “one new species every 24 weeks since 1990” (Cohen and Carlton 1995). Many ecosystems have been subjected to introductions and invasions, although attention to estuaries has been relatively recent compared to freshwater and terrestrial ecosystems (Ruiz et al. 1997). Introduced and invasive species are a global problem (Lodge et al. 2006). There have been about 50,000 nonnative species introduced into the United States thus far; some have been beneficial.
The many species invasions into the bay and delta are not a new problem, but their effects on the ecosystem seem to be increasing. Before 1870, most nonnative species arrived as fouling organisms attached to ships, which is no longer a major problem. Striped bass, which include delta smelt and juvenile salmonids in their diet, were brought by train from the eastern United States and deliberately introduced in the 1870s (Lampman 1946). From 1870 to the early 1900s, other Atlantic species were brought from eastern North America by train and planted in San Francisco Bay; they include oysters, American eels (Anguilla rostrata), lobsters (Homarus americanus), and American shad (Alosa sapidissima) (Lampman 1946).
Although the oysters, lobsters, and eels did not become established, many nonnative species associated with the oysters did become established (Ruiz et al. 1997). Since then, nonnative species have been largely introduced from ballast water. There was a rapid increase in introduced species beginning in the 1940s when ships converted from dry ballast to wet ballast (Thompson 2005). Some invasive10 species arrived, appeared to be increasing, and have since disappeared; Sinocalanus spp. was introduced in 1978 and reached high levels (Orsi et al. 1983) but had dropped in abundance by about 1990 (Winder and Jassby 2011).
Other invasive species have persisted and some have become dominant in the bay and delta. Nearly all common macroinvertebrates present in inner shallows of the bay are introduced species (Nichols et al. 1986). The Asiatic freshwater clam is prevalent in the freshwater areas of the upper delta (Jassby 2008), and the green crab (Carcinus maenas) invaded the bay in 1989-1990 (Cohen et al. 1995). Other examples include Brazilian waterweed, whose areal coverage increased more than 10 percent per year from 2004 to 2006 (Baxter et al. 2010), and largemouth bass (Micropterus salmoides), whose abundance followed that of Brazilian waterweed (Brown and Michniuk 2007). In addition, the frequency of Microcystis aeruginosa blooms (native to the bay and delta) have increased since 1999, concentrated in the freshwater of the central delta during summer (Lehman 2010), and also have affected community composition.
A series of studies has documented the high degree of establishment of nonnative species within the fish community. Feyrer (2004) examined larval fish composition during 1990-1995 in the south delta region and captured 15 species or taxonomic groups, with 3 comprising 98 percent of the total catch by number. The three most abundant species were the alien Asian shimofuri goby (Tridentiger bifasciatus) (71 percent), nonnative eastern and central U.S. threadfin shad (15 percent), and the native prickly sculpin (Cottus asper) (12 percent). Grimaldo et al. (2004) also sampled fish larvae but in four marsh sites in the central delta. They also found that nonnative species dominated the catch, with threadfin shad, members of the sunfish family (Centrarchidae), and inland silversides (Menidia beryllina11) accounting for about 60 percent of the catch. They suggested that the extensive colonization by the nonnative Brazilian waterweed provided good habitat for Centrarchidae (fishes of the sunfish family). Brown and May (2006) examined juvenile and adult fishes through the Sacramento–San Joaquin Delta and found that the overall catch was 59 percent nonnative
![]()
10 “Invasive” species are nonnative species that not only become established but become major components of the ecosystem.
11 This species is designated by some as M. audens, the Mississippi silverside, but we follow AFS (2004) here, as in other fish names.
species, with 93 percent nonnative in the San Joaquin River and 89 percent in the interior delta.
Corbicula amurensis is an example of an invasive species of clam that subsequently caused major shifts in the bay-delta ecosystem. These shifts then act as stressors on the listed fish species. C. amurensis spread in the delta after its introduction in 1986. Nichols et al. (1990) documented how in Suisun Bay the arrival of the clam was correlated with the loss of the dry-period benthic community, despite periods of low flow since the invasion. Winder and Jassby (2011) described how since the invasion chlorophyll a decreased in Suisun Bay and shifted from diatoms to a higher proportion of chlorophytes, flagellates, and cyanobacteria.
The effects of C. amurensis on zooplankton and fish were not as clear as their effects on benthos and phytoplankton. Zooplankton biomass generally declined in the area from Suisun Bay to the central delta over the 1972-2008 period, with some suggestion of declines in particular zooplankton taxa in the delta subregion during the 1980s with the arrival of C. amurensis and an extended drought period (Winder and Jassby 2011). Kimmerer (2002) performed a similar analysis as Winder and Jassby but focused on certain key zooplankton taxa and also included fish. He examined the effects of flow as well, contrasting before and after the C. amurensis invasion. Chlorophyll a decreased between before and after C. amurensis, and there were species substitutions within the zooplankton that offset species-specific losses and thus dampened the decrease at the total biomass level. Pseudodiaptomus affinis replaced Eurytemora affinis, and introduced mysids partially offset the loss of Neomysis mercedis. Despite changes in zooplankton, striped bass survival was not related to the appearance of C. amurensis. Kimmerer (2006) further analyzed an expanded version of the data and suggested that the summer decline in northern anchovy in the low-salinity region was due to their movement out of the area in response to lowered food availability. Diets of other fish species have also responded to the invasion of C. amurensis (Feyrer et al. 2003, Nobriga and Feyrer 2008, Grimaldo et al. 2009a). How these changes in zooplankton composition and diet, and displacement to other areas, have affected fish at the population level is difficult to quantify.
Recently, Glibert and colleagues (Glibert 2010, Glibert et al. 2011) analyzed the long-term data and concluded that changes in nitrogen (concentrations and ratios) were also coincident with some of the changes in chlorophyll a and some key zooplankton species such as E. affinis. They interpreted their results as being a more consistent explanation in terms of timing of declines than the invasion of C. amurensis. However, the matter is not settled (e.g., Cloern et al. 2012). Nonetheless, the analysis of Glibert and her colleagues illustrates the difficulties in attributing dynamics in a
complex food web to single stressors, such as species invasions or changes in a single nutrient.
Conclusions
There is no doubt that nonnative species have affected delta smelt and other fish species listed under the Endangered Species Act. The changes in habitat (e.g., spread of Brazilian waterweed), zooplankton biomass and composition, and predator mix and abundances (e.g., striped bass and largemouth bass are piscivores) have been dramatic, and it is intuitive to look at these changes and infer that such large changes must have had effects at the population level of the fish species. However, such arguments are insufficient for conclusive statements because of the complexity of the linkages between population responses and changes in habitat, food, and predation (Rose 2000). At present, we cannot determine the magnitude of these effects because the relationships among invasives, other stressors, and the listed fish species population responses are complex. Some nonnative species have been present in the ecosystem for more than a century. Some species invasions were localized regionally, preventing easy extrapolation to the fish population level. Also, the invasive species can interact with other stressors, which also are affected by other factors than invasives. The linkage between introduced species and fish species of interest is often due to physical alterations of habitat, shifted food base, or changed predation pressure, and we lack the data or models to make these linkages quantitative. Several analyses have included covariates related to introduced species in the analysis of POD species declines but without definitive conclusions (discussed further below).
Nonnative species as a stressor will continue into the future and likely will become a more prevalent issue. There will be increasing human population and more shipping traffic. Overlaid on these trends are the possibility of large-scale levee failures, sea level rise, and climate change altering the ecosystem and creating new opportunities for invasive species (Moyle 2008).
Nonnative species constitute a stressor that is mostly beyond the control of humans. Prevention is the key but prevention is expensive, requires extensive local, national, and international cooperation, and is risk based. Most introduced species do not become established, but even preventing 95 percent of potential invaders from arriving might be insufficient, because 5 percent could be enough for sufficient inoculations to lead to an invasion. Most legal instruments focus on preventing introductions (Williams and Grosholz 2008). Eradication of some plant species, once they have invaded, is possible, but controlling aquatic animal species, especially mobile species, is not practical. Williams and Grosholz (2008) argue that it is feasible
to control invasive species in marine systems. Interestingly, their examples of successful eradication were plants or generally sessile organisms; no zooplankton or fish examples were given. As inexpensive and convenient control measures become available, they should be evaluated. One example is the use of nitrogen gas to kill organisms in ballast water, which is inexpensive and has the additional benefit of reducing corrosion (Tamburri et al. 2002). But for the most part, introductions and invasions will continue to be a feature of the bay-delta ecosystem and likely will interact with existing stressors (e.g., further changes in the zooplankton community) and might lead to the development of new stressors (e.g., disease).
Early detection is critical, because even if we cannot control the outcome, we can at least make adjustments in monitoring and prepare for possible ecological effects. Introductions will continue, and it is very likely that some of these will lead to successful establishment. Furthermore, also it is likely that a few of these establishments will significantly alter the ecosystem. Such changes to the ecosystem can put endangered species at additional risk and reduce or eliminate the positive effects of management actions. Preparation should involve identifying the likely types of invaders and their possible effects on the ecosystem. Methods exist for identifying vulnerabilities to invasions in ecosystems (Lodge et al. 2006), and the long history of introductions and successful invasions of the bay-delta ecosystem can provide a test bed for evaluating the various vulnerability models. If the possible types of likely invaders and subsequent alterations to the bay-delta ecosystem alterations can be grouped and generalized with some confidence, then some form of contingency planning might be appropriate. Planning can include changes to the monitoring program to allow for earlier detection, and adjustments to planned management actions to prepare for possible ecological effects if such changes occur.
Toxic Chemical Contamination
Chemical contamination is recognized in all plans for the future of the bay delta as a stressor, one of the threats to native and listed species and a factor in regional-scale ecological changes (Healey et al. 2008b). Contamination is not a single issue. There are many contaminants, many of which pose risks to different species, in different locations, or at different spatial and temporal scales. Chemical contamination is historically well documented in the San Francisco Bay Delta compared to many coastal environments (e.g., Luoma and Phillips 1988, van Geen and Luoma 1999, Hunt et al. 2001, Kuivila et al. 2008, Weston and Lydy 2010, Davis et al. 2011). Conceptual models describing processes important in the ultimate impacts of some aspects of chemical contamination were developed for the Delta Regional Ecosystem Restoration Implementation Plan (DRERIP)
process (see p. 128). Nevertheless, recent reviews of the pelagic organism decline concluded either that data are insufficient to demonstrate whether contaminants have adverse impacts in nature (Johnson et al. 2010) or that “ecological effects of contaminants remain unquantified, and are difficult to investigate with standard methods based on acute toxicity” (Brooks et al. 2012).
One problem is that the complexities of the responses of individuals and populations to contamination make it difficult to unambiguously link environmental contamination to specific ecological responses (Luoma and Rainbow 2008, Brooks et al. 2012). Powerful technologies exist to effectively determine concentrations of many potentially toxic chemicals in nature. But because of the limitations of toxicity testing, the complexities of chemical behavior in the environment, and complexities of biological responses, it is difficult to predict with accuracy the concentration thresholds at which local sensitive species will begin to disappear in nature.
A complex combination of considerations determine if chemical contamination is going to be influential in nature:
• the specific chemical’s toxicity, persistence, and tendency to enter food webs;
• the concentration and interactions of that chemical in the environment;
• the spatial scale over which concentrations of contaminants are elevated;
○ many localized hot spots can be as important as region-wide contamination;
• the risks to communities and ecosystem functioning, as determined by differences among species in their
○ physiological tolerance,
○ exposure as determined by functional ecology,
○ genetic flexibility,
○ demographic plasticity, and
○ role in the community (keystone species or important prey species); and
• the time it takes for chronic exposures and subtle effects to manifest themselves as ecological change.
Another issue is that studies of contaminants in nature that include adequate chemistry, biology, and ecology to evaluate impacts are difficult, rare, and considered inadequate evidence by some (Luoma and Rainbow
2008). Fragmented regulatory approaches (see Chapter 5) and important fundamental differences among research disciplines contribute to a lack of synthesis between ecology and ecotoxicology (Luoma and Rainbow 2008). For example, correlative analyses designed to address causes of the most dramatic ecological changes or change points in the bay delta did not even consider toxic chemicals (Dugdale et al. 2007, Sommer et al. 2007, Thomson et al. 2010). Thomson et al. (2010) state that “[c]ontaminants are too numerous and dispersed, and effects too sporadic and subtle, for any monitoring program to provide useful information for correlative analyses. Thus, these effects must be investigated through more detailed, mechanistic studies.”
Despite these challenges, San Francisco Bay is also one of only a few estuarine locations where site-specific ecological impacts from contaminants have been clearly shown in the field. Most obvious is the general observation that since the 1980s, visible impacts of contamination have declined along with concentrations of chemicals in the environment (as the Clean Water Act was implemented). Fish kills that occurred almost once per day in the bay and its tributaries in the 1980s, despite a lack of eutrophication, are now rare (Luoma and Cloern 1982, Brooks et al. 2012). Top predators (e.g., striped bass), which once commonly contained lesions consistent with organic contaminant effects (Luoma and Cloern 1982), have recovered their health. Finally, spatially broad detection of toxicity in standard toxicity tests in the waters of the delta and the major rivers are less frequent than earlier.
Populations of piscivorous birds that were near local extinction because of reproductive failures are recovering. Several more-specific studies meet the criteria for demonstrating cause and effect with reasonable certainty, including minimization of confounding variables (Brown et al. 2003). Long-term studies of the metals silver and copper began when contaminant concentrations were elevated in the 1970s and followed recovery of benthic species and the associated community as metal concentrations in the organisms declined into the 1990s (Hornberger et al. 2000, Brown et al. 2003). Potentially toxic tissue concentrations of selenium in predatory fish and birds were linked with controlled studies of toxicity to show why selenium affects reproduction in benthic rather than pelagic food webs, and that important benthic predators in San Francisco have sufficient exposure to selenium to produce such effects (Stewart et al. 2004, Presser and Luoma 2006). A well-designed ecological study of multiple stressors showed how mercury impacts reduce shorebird reproduction (Schwarzbach et al. 2006). Careful field applications of sensitive in situ toxicity tests showed that pyrethroid pesticides affect the benthos of stream ecosystems (Weston and Lydy 2010). They also demonstrated the frequency of potential contaminant stress on benthos from contaminated sediments in the bay (Hunt et al. 2001). Careful use of control sites and test sites also showed how polychlorinated biphenyls (PCBs) continue to affect benthic communities (and
probably their predators) at an area of high PCB concentration in the bay (Janssen et al. 2011).
These lines of evidence suggest that toxic chemicals, at least at concentrations typical of the 1980s and before, affected individuals and populations of some species, and probably the structure of some communities in the bay delta. Although it is more difficult to identify ongoing effects, it is reasonable to assume recovery from the past is not complete where chemical contamination has declined but not returned to background levels, and that contaminants cannot be eliminated as a stressor of some influence.
It is difficult to rank the importance of contaminants compared to other stressors for the reasons already discussed. But it is possible to be more specific about how contaminant impacts might differ among themselves. If we use specific criteria as defining risk, it is possible to evaluate the degree of that risk, and what contaminant, organism, locality, environmental condition, or season that risk applies to. Similarly, using defined criteria and mechanistic understanding, it is possible to compare how different groups of organisms might respond to different types of contamination risks.
Thus, risks from contaminants must be considered chemical by chemical, with attention paid to the species at risk and the distribution of the contamination. Although it sounds complex, recognizing this principle actually simplifies conclusions about contaminants. Table 3-2, for example, compares risks among different groups of contaminants. Concentrations, toxicity, bioaccumulation potential, spatial distribution, and trends are used as criteria to define the most important issues.
Trends are used as a criterion because the future contamination issues include those that have not been at least partly solved by historical approaches to remediation. Industrial relocation and large investments in waste treatment during the past four decades reduced, but did not eliminate, some of the most serious sources of toxic contamination from the bay delta (e.g., see special issue of Marine Chemistry edited by van Geen and Luoma (1999); Squire et al. 2002). While some areas with high concentrations of contamination remain (e.g., Janssen et al. 2011), the number of such problems is also reduced. But risks are not declining for some contaminants and, for others, risks could increase or trends are not understood (Table 3-2). Spatial criteria are used because contaminants affecting only a few areas of high concentration create less ecological risk to the system than contaminants with a wider geographical influence. Using these criteria and the perspective of chemical class, Table 3-2 indicates the highest risks to the bay-delta ecosystem are posed by selenium, mercury, and pesticides.
In the 1980s, deaths and deformations in birds, along with the local extirpation of aquatic species, accompanied the disposal of selenium-rich irrigation drainage from the western San Joaquin Valley into the Kesterson National Wildlife Refuge. The linkage between selenium contamination and
TABLE 3-2 Contaminants That Have the Greatest Potential for Risks to Bay-Delta Ecosystems as Determined from Their Concentrations, Toxicity, Bioaccumulation Potential, and Trendsa
| Contaminant | Trends | Location | Potential measures |
| Seleniumb | No trend. Potential upward because of high potential for further inputs from the western San Joaquin Valley. | San Joaquin River through Suisun and San Pablo bays. Effects on sturgeon and waterfowl in Suisun/San Pablo bays | In-valley solutions in western San Joaquin Vally. Consider San Joaquin River inputs to bay when evaluating infrastructure changes. |
| Methylmercuryb | No trend. Potential upward if marsh restoration exacerbates methylation. | bay-delta-wide. Effects on birds in South Bay. | Control Hg methylation potential in restored wetlands. |
| Pesticides/ herbicidesc | Unknown. High usage continues. | Worst effects in local sloughs and urban streams and rivers. Enough stress points to make this a regional problem? Pesticide squeeze. | Best management practices (orchard pesticide example; Werner et al. 2004). |
| Emerging chemicals (pharmaceuticals, etc.)c | Upward? Little spatial information. | Localities influenced by poorly treated urban wastes. | Waste treatment. |
| Metals (Ag, Cd, Cu, Pb, Zn, V, Ni, Cr)d | Downward: 1970-2000. Stable recently. | Urbanized areas. Mine impacted areas upstream. Perhaps delta islands where Cu is in herbicides. | Sustain point source waste treatment. Remediate mine wastes impacts. |
| Legacy organic contaminants and PAHsd | Downward. No trend for PAHs. | Urbanized areas. | Clean up legacy hot spots, especially in bay. |
aLocations and food webs at risk are also shown (these differ among contaminants), as are potential measures for managing these risks.
bHigh certainty that this is an important stressor with potential for increased problems in the future. Long-term need for increased management.
cPockets of contamination exist with high certainty of adverse ecological impacts. Uncertainty as to whether enough stress points exist in time and space to make this a regional-scale stress. Need for long-term improved management is certain.
dTemporal trends show these potential stressors have declined in recent decades, although concentrations of most remain moderately elevated. Sustained management is essential.
NOTE: PAH, polycyclic aromatic hydrocarbon.
toxicity to wildlife was unambiguous at the Kesterson National Wildlife Refuge.
A very large reservoir of selenium exists in the soils of the western San Joaquin Valley associated with the salts that accumulated there during decades of irrigation (Presser 1994). Irrigation drainage, contaminated by selenium from those soils, is also accumulating in western San Joaquin Valley groundwaters. The problem is exacerbated by the recycling of the San Joaquin River when water is exported from the delta. While control of selenium releases into the San Joaquin River from the valley soils has improved, how long those controls will be effective is not clear because of the selenium reservoir in groundwater.
Some potential solutions could create more problems than they solve. For example, proposals to dispose of the contaminant outside the San Joaquin Valley in the bay or in the oceans could exacerbate ecological risks there (Presser and Luoma 2006). Other aspects of water management also could affect selenium contamination. For example, infrastructure changes in the delta such as construction of an isolated facility could result in the export of more Sacramento River water to the south, which would allow more selenium-rich San Joaquin River water to enter the bay. The solutions to selenium contamination must be found within the Central Valley and the risks from selenium to the bay are an important consideration in any infrastructure changes that affect how San Joaquin River water gets to the bay.
Organochlorine pesticides like DDT were unquestionably a cause for the near extirpation of piscivorous bird populations in the bay delta in the 1970s and 1980s. More recently, pesticide toxicity that was once dominated by water column effects attributable to pesticides like carbamates has switched to contaminated sediments as the dominant class of pesticides has switched to pyrethroids (Weston et al. 2005). Benthic food webs dependent for a part of their life cycle on urban streams, sloughs, as well as floodplains, and streams or rivers that receive direct runoff from cities or agricultural fields, appear to be at risk from the growing use of this class of pesticide. Because sediment-bound pesticides enter aquatic systems with the high sediment concentrations that accompany the first flush of agricultural fields and urban landscapes, species that are mobilized during such a period (e.g., delta smelt) may also be more at risk. The sensitivity to pyrethroids of native species, the spatial distribution of the contamination, its seasonality, its food-web dynamics, and effects on community structure and function are not as well known as they need to be. But pesticides are an important stressor in at least some localities.
Concerns about mercury stem from a historic legacy of widespread mercury contamination north of the delta (Suchanek et al. 2008, Bouse et al. 2010); efficient biomagnification of methylmercury in food webs; high toxicity of methylmercury to reproduction of upper-trophic-level species;
threats to the health of people that consume certain species of fish from the watershed (Greenfield et al. 2005); and the possibility that restoration of wetlands could exacerbate the methylation of mercury in sediments.
Green sturgeon (Acipenser medirostris) appears to be the species most at risk from chemical contamination. Sturgeon tissues contain higher concentrations of selenium and mercury than any other fish species, reflecting their position as a top predator in the benthic food web (Stewart et al. 2004). This may also result in greater exposure to bioaccumulative organic contaminants, such as PCBs and perhaps some emerging chemicals of concern. Because green sturgeon is a long-lived, slowly reproducing species, populations are vulnerable to chemical disruption of reproductive processes (typical effects of selenium and mercury). The few analyses of sturgeon populations consistently fail to mention contaminants in the list of sturgeon stressors; an illustration of the scientific disconnect between ecology and ecotoxicology (Luoma and Rainbow 2008).
Risks from mercury provide an example of the complexity of ranking contaminants as a stressor. Schwarzbach et al. (2006) showed that mercury contamination exacerbated low reproduction potential in the endangered California clapper rail (Rallus longirostris), a shore bird in the south bay. Loss of habitat is the most important stressor for the clapper rail in the bay. Within the existing habitat, however, Schwarzbach et al. (2006) first considered how nests of the species were affected by predation and flooding. After those effects were accounted for, the nests most contaminated with mercury had the lowest reproductive success rate. Thus, mercury is not the only stressor for clapper rails, but it is one of the stressors holding back recovery of this endangered species. Runoff into South Bay from the historic New Almaden mercury mine is the source of contamination in this ecosystem. More important, a large area of wetlands undergoing restoration in the south bay receives freshwater from the stream that drains the catchment containing this mine. Because methylmercury production is amplified in wetlands, and at least some historic sources continue to release mercury (Suchanek et al. 2008), adding wetland habitat could result in an expansion of the mercury problem in the delta.
Conclusions
Contaminants are not a single ubiquitous stressor in the delta as much as they pose risks that differ among the chemicals, among species, among locations, and among seasons or even years. In no case is it clear that “contaminants” are the sole cause of large-scale ecological change in the delta at present. On the other hand, contaminant stress was likely an important factor in piscivorous birds and benthic communities near outfalls (Hornberger et al. 1999) and fish that were resident in urban streams before
the 1990s (Luoma and Cloern 1982). When implementation of the Clean Water Act began to take effect, however, at least some of the most concentrated contamination was reduced and some of these effects were reversed (Hornberger et al. 1999). In addition, contaminants cannot be eliminated as one of the several causes of some of the ongoing changes in today’s delta like the apparent continuing decline of white sturgeon populations, poor reproduction in certain shore birds, and simplification of benthic communities in streams affected by urban runoff. In addition, it is not clear that sustainable solutions are in place to reduce the effects of contaminants like selenium, mercury, and pesticides and some proposed changes could even increase risks. Nor are there sufficient data to fully understand the implications of some new classes of emerging contaminants. Ranking contaminants relative to other stressors will vary with the perspective of the ranking body. Given the complexities described above, that is probably not a constructive exercise. However, continuing to better understand and address the most important contaminant issues should remain one of the priorities in managing the delta ecosystem.
Impediments to Fish Passage
Impediments to fish passage take a variety of forms. NMFS (2009a,b) applies the term in a broad sense to include structures and actions that can interfere with fish movement through a migratory corridor. This can include dams, unscreened water diversions or pump intakes, and a variety of anthropogenic actions that can produce thermal barriers or other water-quality problems. For this discussion we define passage impediments as structures (e.g., dams) and actions (e.g., diversion of water and pumping facilities) that block or remove fish from the migratory corridors upstream from the delta. Impediments have a range of effects, from slowing the migration by delaying passage, removing fish from the migration corridor to encounter hazardous conditions, or completely blocking access to productive habitat.
Dams as Absolute Barriers
One prevalent form of passage impediment in the Central Valley is dams that form absolute barriers to migrating fish, in that they have neither ladders for adults nor bypass systems to pass seaward-bound smolts. Dams have been built for a variety of purposes, including hydropower, flood control, irrigation, and municipal uses. Many have permanently blocked or hindered salmonid access to historically productive spawning and rearing grounds and have dramatically truncated the freshwater habitat accessible to anadromous salmonids and sturgeon. These impacts were evident more
than 80 years ago when Clark (1929) estimated that 80 percent of this habitat for these species in the Central Valley had been lost by 1928. More recently Yoshiyama et al. (1996) estimated that 82 percent of the historical salmon habitat is now inaccessible. NMFS (2009b) suggests that the extent of habitat loss for steelhead (Oncorhynchus mykiss) may be even greater, since they had a broader geographic distribution than Chinook salmon.
As a result, winter-run and spring-run Chinook salmon, and steelhead populations, are confined to lower-elevation portions of many tributaries as well as the mainstem Sacramento and San Joaquin rivers (NMFS 2009a, b). Overall this decrease in the quantity and quality of spawning and rearing habitats has reduced fish abundance (Lindley et al. 2009). The reduction of a habitat type not only limits potential carrying capacity but also negatively affects the population structure of anadromous fish, by reducing the number of independent population units. Lindley et al. (2004) note that only one population of winter-run Chinook now exists, restricted to a confined temperature-regulated zone below Keswick Dam. They suggest that historically four separate populations inhabited the Central Valley. Spring-run Chinook salmon have incurred the same fate, with only 3 of 19 historical independent populations remaining. Lindley et al. (2006) estimate that no fewer than 81 independent steelhead populations once existed in the Central Valley.
The southern distinct population segment (DPS)12 of green sturgeon incurred a similar fate. One population is currently confined to a single spawning area in the upper mainstem Sacramento River. Historically spawning habitat likely extended upstream from the current site into the Little Sacramento, Pitt, and McCloud rivers (Adams et al. 2007). Green sturgeon may also have spawned in the Feather River, upstream from Oroville Dam.
Viable Salmonid Populations (VSP)
The reduction in population complexity associated with migratory barriers affects not only fish abundance by limiting the quantity of suitable habitat but also ultimately the probability of the species persisting in the Central Valley. The Viable Salmonid Populations (VSP) framework (McElhany et al. 2000) provides a foundation for discussing these impacts. The VSP parameters of productivity, abundance, and population spatial
![]()
12 The Endangered Species Act (ESA) defines the term “species” as including “any subspecies of fish or wildlife or plants, and any distinct population segment of vertebrate fish or wildlife which interbreeds when mature” (Section 3 (15)). A DPS is thus a smaller evolutionary unit than a species or subspecies. (If a DPS is the whole species, then it is called a species and not a DPS.) For more detailed discussion of this term, see NRC (1995).
structure are key indicators of a species’ resilience and likely viability. Reduction in the values of these parameters is associated with a loss in genetic or life history variability. Ultimately this results in reduced population resilience to environmental variation at local and basin-wide scales.
The committee concludes that the dams that act as absolute barriers, which have eliminated access to nearly 80 percent of the historical habitat, have been, and continue to be, a major stressor limiting the recovery of ESA-listed anadromous fish species in the Central Valley. The effects include limiting abundance and productivity associated with severe habitat loss, and the pronounced reduction of genetic diversity through extirpation of the vast majority of unique populations once present in the system (e.g., NRC 1996).
Dams as Partial Barriers
The Red Bluff Diversion Dam (RBDD) is owned and operated by the Bureau of Reclamation. It is located 59 miles downstream of Keswick Dam. For decades until 2011, the dam blocked or delayed adult salmonids and sturgeon migrating upstream to various degrees, depending on run timing and configuration of the dam during the different migratory periods (CDFG 1998, Vogel et al. 1988). Dam operations affect both juvenile and adult life stages of salmonids, and sturgeon. The intent is that after May 2012 the gates will be permanently opened and irrigation water will be provided by pumps (USBR 2011).
The Anderson-Cottonwood Irrigation District (ACID) diversion dam spans the Sacramento River 5 miles downstream from Keswick Dam. It is one of the three largest diversions on the Sacramento River, and the ACID has senior water rights of 128,000 acre-feet of water. The diversion dam is operated from April through October. Substantial reductions in water releases from Keswick Dam are required to install or remove the flashboards at the dam. This operation has dewatered redds, and stranded juveniles. However, the reductions in flows usually last for less than 8 hours, but the amount of mortality due to dewatering of incubating eggs and stranding juveniles is uncertain. Even so, this constitutes a risk to early life stages. Based on run timing, the diversion dam operations could affect winter-run, spring-run, and fall-run Chinook and green sturgeon (Table 3-3).
The ACID diversion dam was improved in 2001 with the addition of new fish ladders and fish screens around the diversion (CDFG 2004). Since upstream passage for salmonids was improved, winter-run Chinook spawning shifted upstream with more than half of the winter-run redds typically observed above the ACID diversion dam. The majority of winter-run in recent years (i.e., > 50 percent since 2007) spawn in the 5 miles of river from Keswick Dam downstream to the ACID dam (NMFS 2009b). Nevertheless,
the ladders do not accommodate green sturgeon, and thus the migration is completely blocked during a portion of the migratory period (Table 3-3). Newly emerged green sturgeon larvae that hatch upstream of the ACID diversion dam would be forced to remain for 6 months upstream of the dam or pass over it and be subjected to higher velocities and turbulent flow below the dam, thus rendering the larvae and juvenile green sturgeon more susceptible to predation.
Given the paucity of quantitative studies of survival probabilities associated with passing or operating seasonally passable dams, we cannot determine the extent to which they have contributed to the decline of the ESA-listed anadromous species in the Central Valley. Even so, we suspect the effect was historically pronounced, has diminished in the past decade, and may diminish further as new operations are developed.
Smaller Water Diversions
Apart from the larger dams that span the mainstem or major tributaries, a complex of smaller water diversions for irrigated agriculture, municipal and industrial use, and managed wetlands are found throughout the Central Valley. According to NMFS (2009a), thousands of small and medium-size water diversions exist along the Sacramento River, San Joaquin River, and their tributaries. Many remain unscreened. Herren and Kawasaki (2001) reported that 98.5 percent of the 3,356 diversions included in a Central Valley database were either unscreened or inadequately screened.
Unscreened diversions put juvenile fish at risk by removing them from the rivers, thus contributing to mortality during their rearing phase or seaward migration. Although actual entrainment rates are not cited, NMFS (2009a) states that the CVP/SWP operations Biological Assessment (USBR 2008) provided calculations of estimated entrainment of salmonids through unscreened diversions along the Sacramento River. According to the calculations, over 7,000 juvenile winter-run are lost to unscreened diversions annually. Beyond this we have not encountered reliable estimates of population-level effects on any anadromous species, as associated with entrainment by water diversions. NMFS (2009a) notes that estimates of the mortality at unscreened diversions in the Sacramento River are small, but the cumulative impact is likely to affect ESA-listed species at the population level. NMFS (2009a) also notes that most, but not all, large diversions are screened. To guide future fish screening projects, the Anadromous Fish Passage Program is focusing on monitoring losses at smaller unscreened diversion. NMFS (2009a) concludes that the combined mortality from all screened diversions in the Sacramento River is insignificant at the population level for the ESA-listed species. However, Moyle and Israel (2005) note the paucity of information regarding population-level effects of diver-
TABLE 3-3 Life History Timing for Anadromous Fish Species in the Upper Sacramento River
| Species | Adult Immigration |
Adult Holding |
Typical Spawning |
Egg Incubation |
Juvenile Rearing |
Juvenile Emigration |
| Winter-run Chinook | Dec – Jul | Jan – May | Apr – Aug | Apr – Oct | Jul – Mar | Jul – Mar |
| Spring-run Chinook | Apr – Jul | May – Sept | Aug – Oct | Aug – Dec | Oct – Apr | Oct – May |
| Fall-run Chinook | Jul – Dec | n/a | Oct – Dec | Oct – Mar | Dec – Jun | Dec – Jul |
| Late fall-run Chinook | Oct – Apr | n/a | Jan – Apr | Jan – Jun | Apr – Nov | Apr – Dec |
| Steelhead Chinook | Aug – Mar | Sept – Dec | Dec – Apr | Dec – Jun | Year round | Jan – Oct |
| Green sturgeon Chinook | Feb – Jun | Jun – Nov | Mar – Jul | Apr – Jun | May – Aug | May – Dec |
SOURCE: Reproduced from Table 5-1 in NMFS (2009a).
sions in the Central Valley and conclude that screen diversions may have population-level effects.
The weight of evidence—or in this case lack thereof—indicates that the impacts of screened and unscreened water diversion on anadromous fish are poorly described and certainly not quantified in any meaningful manner. Given this, the contribution of this class of stressors to the decline of anadromous fish in the Central Valley is unknown. A thorough evaluation of water diversions within the active migratory corridor is warranted.
Delta Pumps and Related Flow Effects
The committee fully appreciates the complexity of mechanisms and negative impacts that the SWP and CVP pumping operations have on juvenile salmonids in the vicinity of the delta. The National Research Council’s (NRC’s) 2010 report on the delta (NRC 2010) noted that, in addition to direct effects associated with entrainment at the pumps, there are indirect effects associated with predation within the labyrinth of delta channels. The dynamics is further complicated by magnitude and timing of OMR flow (NRC 2010).
The committee accepts the conclusion that pump operations pose a risk to juvenile salmonids. The survival of salmonid smolts migrating through the delta is low. Several studies make this point. Recently, Michel (2010) used acoustic-tagged late-fall Chinook yearling smolts to estimate survival from the upper Sacramento River (Battle Creek) to the mouth of San Francisco Bay. Expressing survival in each segment in terms of survival per 10 km of migration distance, he found low survival in the upper Sacramento River from the release site to near Butte City (92.4 to 96.8 percent/10 km) and through the delta zone (93.7 percent/10 km). The lowest survival occurred through the San Francisco Bay estuary immediately west of the delta (67.0 to 90.2 percent/10 km). Based on these results, survival of yearling Chinook salmon through the delta is estimated to be 52.5 percent (±3 S.E). In support of this estimate, Perry et al. (2010) reported delta survivals of Coleman hatchery-origin late fall–run Chinook salmon smolts of 35 percent (±10 S.E.) and 54 percent (±7 S.E.) in December 2006 and January 2007, respectively. In contrast, Michel estimated total survival from release in the upper Sacramento River to the mouth of the San Francisco Bay was an order of magnitude lower, ranging from 3.1 to 5.5 percent. Michel also noted that this total survival (which includes the delta segment) was substantially lower than published values for other west coast yearling Chinook. Notably, it is an order of magnitude less than that typically reported for yearling Chinook smolts migrating past eight dams in the Snake Columbia River system.
It was not possible to ascertain from these data the magnitude of direct
and indirect effects associated with pump operations as smolts migrated through the delta. Nevertheless, visual inspection of the survivorship curve by Michel (2010) suggests that on average perhaps 20 to 30 percent of the smolts died while migrating through the delta zone as delineated in that study. These losses are substantive and are at least in part attributable to pump operations that alter current patterns into and through the channel complex, drawing smolts into the interior waterways and toward the pumps.
Statistical analysis of tagged hatchery releases recovered at Chipps Island or the ocean fishery have shown negative associations between pump export volume and relative survival. However, the variation in relative survival was very large (Newman and Rice 2002; Newman 2003, 2008).
The mortality of smolts migrating out of the San Joaquin River drainage and through the delta is also pronounced. Recent studies using San Joaquin River fall Chinook salmon smolts estimated survival between 5 and 8 percent as smolts migrated through the south delta, Old River, and reaches leading to the pumps (San Joaquin River Group Authority 2010). Furthermore, preliminary survival information has suggested that San Joaquin fish collected at the south delta pumps and transported out of the delta had higher, but still very low, survival than fish that migrated through the San Joaquin River (R. Buchannan, personal communication to J. Anderson). The committee recognizes that these estimates are for one salmon species only and others may exhibit different responses. However, at this juncture these estimates provide the best available population-level index of impacts associated with passage past and through the delta during periods of pump operations.
Strategies for mitigating the impacts of mortality of juvenile salmon passage through the delta are likely to differ for Sacramento and San Joaquin runs because of the differing routes through the delta. The Sacramento fish can avoid the higher mortality of the central delta altogether by entering the Yolo Bypass when it floods (Figure 3-7, ![]() ) or by passing through the lower Sacramento when the Delta Cross Channel is closed (Figure 3-7,
) or by passing through the lower Sacramento when the Delta Cross Channel is closed (Figure 3-7, ![]() ). In contrast, juvenile fish migrating through the San Joaquin are either routed directly through the delta (Figure 3-7,
). In contrast, juvenile fish migrating through the San Joaquin are either routed directly through the delta (Figure 3-7, ![]() ) or toward the south delta pumps (Figure 3-7,). It has been generally believed that routing fish away from the pumps is undesirable (route
) or toward the south delta pumps (Figure 3-7,). It has been generally believed that routing fish away from the pumps is undesirable (route ![]() preferred over
preferred over ![]() ), which has been the main justification for closing the barrier at the head of the Old River. However, recent studies have suggested that survival through Old River with collection at the pumps and transport out of the delta may provide better, although still low, survival than when routing fish through the main channel (route
), which has been the main justification for closing the barrier at the head of the Old River. However, recent studies have suggested that survival through Old River with collection at the pumps and transport out of the delta may provide better, although still low, survival than when routing fish through the main channel (route ![]() ). Possible reasons for this surprising conjecture may involve differences in predator densities in the routes as well as differences in tidal influences on passage. Juveniles may experience multiple encounters
). Possible reasons for this surprising conjecture may involve differences in predator densities in the routes as well as differences in tidal influences on passage. Juveniles may experience multiple encounters

FIGURE 3-7 Alternative routes of passage of Sacramento and San Joaquin salmon through the delta. Sacramento River fish diverted into Yolo Bypass avoid the central delta ![]() , while those not entering Yolo are susceptible to entering the delta
, while those not entering Yolo are susceptible to entering the delta ![]() . San Joaquin fish either pass through the delta in the main stem of the river
. San Joaquin fish either pass through the delta in the main stem of the river ![]() , or enter the Old River to be drawn to the pumps where the survivors are collected and transported out of the delta
, or enter the Old River to be drawn to the pumps where the survivors are collected and transported out of the delta ![]() .
.
SOURCE: California Natural Resources Agency (2010).
with predators as they migrate through the delta and are displaced back and forth by the tides. In contrast, fish moving with the flow toward the pumps may experience weaker tides and thus fewer multiple encounters with predators (Anderson et al. 2006). The salient point is that survival of fish through the San Joaquin system is complex and further survival studies are needed to evaluate what actions might be most effective in improving their passage through the delta.
The available data based largely on Michel’s (2010) and the San Joaquin River Group Authority (2010) studies suggest that delta-specific management actions may not yield the large survival benefits some might expect. Migrating smolts incur substantial levels of mortality outside of passage through the delta, including mortality directly and indirectly associated with SWP and CVP pump operations. The take of fish at the pumps represents a fraction of the total population that is drawn toward the pumps through the various delta reaches. Mitigating the effects of pumping
involves routing fish thought the delta segments with the lowest mortalities as well as mitigating the take directly at the pumps. Thus, control over fish passage routes and improved collection and transport of salmonids at the pumps both need to be considered as mitigation actions. However, at this time the data and understanding of mortality processes within the delta are insufficient to identify a course of action. Increasing passage through Yolo Bypass may be a viable action for Sacramento runs. However, actions for San Joaquin fish appear less certain. Should actions divert fish through the tidally dominated central delta or should fish be diverted, collected, and transported at the pumps? Information is insufficient to evaluate such alternatives.
Delta Smelt
The entire life cycle of the delta smelt is confined to the delta region, which includes the area where the pumps are situated. Population-level effects of entrainment from pump operations have been described by Kimmerer (2008b), and subsequently critiqued by Miller (2011), which led to a reevaluation by Kimmerer (2011). In the initial Kimmerer (2008b) analysis, estimated overall impacts were generally small to moderate in most years (< 20 percent), but were high (> 30 percent) in some of the years analyzed. Kimmerer (2008b) noted that the estimates have large confidence limits and have index values varied widely across years, with large proportional losses of some delta smelt life stages evident in some years. He suggests that these highly variable annual loss estimates reflect episodic effects and therefore their annual magnitude should be empirically calculated rather than inferred from correlations.
Subsequent to Kimmerer’s (2008b) analysis, Miller (2011) systematically laid out the assumptions and data issues in the Kimmerer (2008b) analysis. He then inferred that because most of the assumptions made by Kimmerer (2008b) would lead to an upward bias in the estimated population-level impacts, Kimmerer’s estimates of impacts were therefore high. Kimmerer (2011), in his reanalysis, addressed some but not all of the issues and uncertainties raised by Miller (2011). Kimmerer (2011) concluded that, while the new estimates were slightly lower, the initial conclusion that entrainment by the pumps was large on an episodic basis remained valid.
The difficulties in estimating population-level impacts from entrainment are illustrated by the many assumptions and the complexity of the analyses detailed in all three of these papers. Continued critiques and constructive exchanges will enable further refinement of the estimated impacts and clear identification of the key uncertainties that need to be addressed with additional modeling and data collection.
Based largely on Kimmerer (2008b, 2011), Miller (2011), and the NRC’s previous conclusion on pumping operations (NRC 2010), we conclude that in some years the population-level impacts on delta smelt are large and thus is a significant factor affecting delta smelt population dynamics. The committee does note that the status and knowledge of delta smelt have changed substantially in the last 5 years. Take at the pumps has been low and the seasonal sampling programs have suggested that the population levels are extremely low. However, an experimental sampling protocol coordinated with tidal cycles recently found unexpected concentrations of adult delta smelt. We therefore conclude that, in general, significant uncertainty exists on the condition and prospects for recovery of the delta smelt.
Model Analyses of Pump and Flow Effects
In an earlier report (NRC 2010), the assessment of the modeling framework required to adequately assess effects among pump and flow treatment noted some significant deficiencies that will impede informed decision making. The report emphasized the need for a more comprehensive life-cycle modeling approach that is more realistic and better matches the scale of processes at the population level (NRC 2010, pp. 40-41). This committee concludes that population-level effects analysis is required in order to rank this class of stressor against the others identified in this report. Absent that solid quantitative perspective, we are left to rely on qualitative assessments (e.g., Delphi process) in ranking entrainment (or any stressor) among all of the possible stressors. We note that this deficiency is not specific to the pump/flow mechanisms in the delta; all the stressors we discuss suffer in this regard.
There is recent accelerating activity in the area of life-cycle modeling of salmon and delta smelt. Several models are under development but not yet published. Maunder and Deriso (2011) recently published a life-cycle model of delta smelt. This model includes some assumptions that need further additional evaluation (e.g., role of density-dependent survival). However, the model is noteworthy because it illustrates that there is increasing activity in the important area of life-cycle modeling. The committee knows of several other life-cycle models that are in various stages of completion and is encouraged by this upsurge in activity. We further encourage continued development of models, within a collaborative regional process. A collaborative process is needed to minimize the paralysis that can occur from dueling models that are difficult to compare after their development and analyses are completed.
Conclusion
The committee concludes that the dams that act as absolute barriers, which have eliminated access to nearly 80 percent of the historical habitat, have been and continue to be a major stressor adversely affecting ESA-listed anadromous fish species in the Central Valley. The effects include drastically limiting abundance and productivity associated with severe habitat truncation, and the pronounced reduction of genetic diversity through extirpation of the vast majority of unique populations once present in the system.
Passage impediments at the RBDD and ACID diversion dam contributed to the decline of the ESA-listed anadromous species in the Central Valley. However, improvements in passage at both facilities, ACID in 2001 and RBDD in 2011, appear to have significantly improved passage in the Sacramento River.
The effects of water diversions on anadromous fish are poorly described, inadequately evaluated, and remain unquantified in any meaningful manner. Given this, the contribution of this class of stressors to the decline of anadromous fish in the Central Valley is unknown. We recommend a thorough evaluation of screened and unscreened water diversions within the active migratory corridor.
Based on smolt survival studies (Michel 2010, SJRGA 2010) and export–flow–survival relationships, the committee concludes that mortality incurred while migrating through the delta is substantial and in part attributable to pump operations. However, we cannot determine the extent to which altering pump operations, or providing alternative passage options, might affect population-level responses (e.g., population growth rate) relative to other stressor agents. The limited studies do indicate that significant mortality occurs prior to smolts reaching the delta pumps. For the delta smelt we conclude that in some years pumping operations pose a high risk to smelt, but in other years the impacts appear low. It is difficult to assess the current impact on the total population because it appears few delta smelt are found in the central delta. On balance we judge that across years this stressor poses a moderate impact to smelt at the population level.
There seems to be an expectation in the region that alleviating or minimizing pump effects in the delta will lead to robust populations of the ESA-listed salmon populations in the Central Valley. Assessing the likely effectiveness of doing this would be helped by developing a comprehensive, life-cycle model that is capable of exploring a variety of passage alternatives in combination with effects from other stressors. Several salmonid models are under development and we encourage their development and cross-comparisons and cross-fertilization. Furthermore, assessing the impacts of pumping on salmon populations will also require further studies to assess the impacts of pumping on the passage routes of smolts through the delta
complex and their survival through the routes. The NRC noted this in an earlier report (NRC 2010).
The committee concludes that an integrated quantitative analysis is fundamental and required in order to rank the SWP and CVP pump operations on fish routing and direct take against the other stressors identified in this report. Absent that solid quantitative perspective we are left to rely on qualitative assessments in ranking among other stressors. This holds not only for anadromous salmonids, but also delta smelt and other species of concern. This deficiency is not peculiar to the pump/flow mechanisms in the delta; many if not all the stressors we discuss suffer in this regard.
Fishing
The potential negative effects of fisheries on individual species (Myers et al. 1996), on ecosystem services (Worm et al. 2006), and on coastal and estuarine ecosystems generally (Jackson et al. 2001) is widely acknowledged. For individual species, fisheries are known to have a range of effects on exploited population beyond the obvious decreases in abundance. Fisheries are highly selective agents of mortality that can cause rapid changes in phenotypic (Rjinsdorp 1993) and genetic traits (Policansky 1993, Conover et al. 2005). Beyond their direct effects on individual species, fisheries can alter community structure (Yemane et al. 2005, Kitchell et al. 2006) and disrupt habitats (Collie et al. 2000). It is not only industrial, commercial fisheries that have the potential to produce these changes; recreational fisheries can also contribute substantially to mortality in exploited species and may influence community structure (NRC 1999, 2006, Ihde et al. 2011). Moreover, even when fisheries occur in restricted geographical regions, their impacts can be felt over a broader geographic range because many species are highly mobile and often undertake long migrations. This is particularly true for estuarine-dependent diadromous species such as the salmonids (e.g., Chinook salmon and steelhead) and temperate sea basses (e.g., striped bass).
The bay and delta supported sizeable fisheries in the past. Historical reports indicate the first commercial fisheries in the bay-delta system developed in the mid-1800s and targeted Chinook salmon in the Sacramento and San Joaquin rivers and in Suisun Bay (Scofield 1956, cited in Smith and Kato 1979). Rapid expansion of these fisheries, in combination with reduced water quality and impediments to stream passage, led to substantial reductions in salmon numbers. However, both commercial and recreational fisheries continued until 2008, when all salmon fisheries in state waters were closed following dramatic declines in Sacramento fall-run Chinook. The closure continued in 2009. It has been estimated that these closures led to economic losses of more than $250 million and more than 2,000
jobs annually (Morse and Manji 2009). Today, the only commercial fisheries that remain in the delta proper are for threadfin shad, armed box crab (Platymera gaudichaudii), and crayfish (CA Fish and Game).13
During this time, commercial fisheries also developed in San Francisco Bay proper. These fisheries targeted a diverse assemblage of species including Pacific herring (Clupea pallasii), striped bass, both white (Acipenser transmontanus) and green sturgeon, and Chinook salmon. Several of these fisheries followed the same pattern of expansion and retraction exhibited in the salmon fisheries (Figure 3-8). Fisheries for other species have had longer histories, including those for anchovy (Engraulis mordax) and Pacific herring. Fisheries for herring harvested as much as 4,000 metric tons (mt) as late as 1975. Although much smaller than at its peak, in 2009, the last year this fishery operated, the California Department of Fish and Game report a harvest of 459 mt landed in the port of San Francisco.14 In many ways the fishery for herring in San Francisco Bay was the last reminder of once sizeable estuarine fisheries. Indeed, inspection of current commercial landings for the Port of San Francisco reveal fisheries dominated by nonestuarine species such as Dungeness crab (Metacarcinus magister) and Pacific sardine (Sardinops sagax).
Today the bay and delta support recreational fisheries for striped bass, largemouth black bass, white sturgeon, Chinook salmon, steelhead, catfishes (family Ictaluridae), and American shad. In 2004, the state of California mandated that any anglers wishing to fish within the estuarine system purchase a Bay-Delta Sport Fishing Enhancement Stamp (California Fish and Game Code § 7361(b)). The funds from this program were invested in research and restoration activities to enhance and sustain recreational fisheries in the region. This program provides the basis for estimates of angler participation. From 2004 to 2009, when the program was closed, 1.81 million anglers purchased more than $9 million worth of stamps (CDFG 2009), suggesting that perhaps 300,000 anglers fished the San Francisco Bay-Delta system annually.
We may identify direct (removal of target species or removal as bycatch) and indirect (removal of potential prey or predators) impacts of fisheries. Elsewhere fisheries have been shown to have important direct (Myers et al. 1996) and indirect effects (Kitchell et al. 2006). Thus, the central question before us is, “Given this pattern of commercial and recreational fisheries, what impact may these fisheries have on the bay-delta ecosystem?”
Considering direct impacts first, the minimum requirement for their assessment are annual estimates of the size of the targeted fish population and an estimate of the total removals by fishing for that population. Often
![]()
13 Available at http://www.dfg.ca.gov/marine/landings09.asp. Accessed May 12, 2011.
14 Available at http://www.dfg.ca.gov/marine/landings09.asp. Accessed May 12, 2011.
these estimates are derived from sampling programs that estimate relative abundance and catch levels, and stock assessment modeling that translates these empirical estimates to absolute impacts. For many species of interest in the bay-delta system, sampling programs are available to estimate trends in abundance (e.g., Fall Mid Water Trawl Survey15) and harvest [e.g., Recreational Fisheries Information Network (RecFIN)16].
However, population models that combine these data streams to provide an integrated picture of trends in absolute abundance and exploitation are lacking for all principal species, a point made also by the NRC (2010). This lack precludes provision of absolute estimates of the impact of harvest on any of the principal species. Despite this shortcoming, it is possible to estimate trends in the relative impact of exploitation on the dynamics of any targeted species.
As an example we consider striped bass in the bay delta and note that similar calculations could be undertaken for other species. The annual salvage estimate at the Tracy Fish Collection Facility (TFCF) may provide an estimate of the relative abundance of striped bass in the system (Aasen 2011; Figure 3-9A). We recognize that the number of striped bass salvaged will be affected by the amount of water conveyed by the system, but as a first approximation inter- and intra-annual variability in the volume of water conveyed will be ignored. It would be equally possible to use other indices of abundance, such as the California Department of Fish and Game tag-recapture. The RecFIN program provides an estimate of the total number of striped bass harvested in inland waters in northern California (Figure 3-9B17). Finally, the ratio of the estimates of harvest and abundance provide a measure of relative exploitation, U (Figure 3-9C). These relative exploitation estimates may provide an indication of the years in which exploitation was relatively more important. For example, the pattern in Figure 3-9C suggests that exploitation was almost twice as large in 1996 and 2003 (U ~ 3500) as in 1993 (U ~ 1700). But importantly, these data cannot be scaled to absolute impacts. Thus, their utility in comparing the importance of exploitation as a stressor among species or among stressors is limited.
Further complicating estimation of relative exploitation rates is the fact that many species of interest in the San Francisco Bay-Delta system are diadromous (migrate between saltwater and freshwater) and have broad distributions. As a consequence, the principal exploitation stress may not be from harvests within the bay-delta system, but rather from harvests outside the system. For example, green sturgeon tagged in the bay-delta
![]()
15 See http://www.dfg.ca.gov/delta/projects.asp?ProjectID=FMWT. Accessed July 16, 2012.
16 See http://www.recfin.org. Accessed July 17, 2012.
17 See http://www.recfin.org/data/estimates/download-estimates-data-files. Accessed July 17, 2012.

system have been recaptured off Oregon. The catch of these species outside the bay-delta system, whether in targeted fisheries such as for salmon or as incidental catch such as the case for green sturgeon, may be an important additional stress.
Estimating the indirect effects of fishing on an ecosystem presents a greater challenge than estimating the direct effects of exploitation. Most often these impacts are observed after the fact rather than forecast a priori (Baum and Myers 2004). Attempts to quantify indirect effects have usually relied on ecosystem-based models of fishery ecosystems using tools such as EcoPath with EcoSim. Such a model is available for the delta system (Bauer 2010). Ecosystem models have a high demand for data, including time series of relative abundance, catch data, and diet information, to quantify the composition and quantity of prey items for each species. Bauer’s model involves 40 functional groups (both trophic groups, individual species, and age classes of individual species). Represented in the model are three age classes of striped bass, Chinook salmon, splittail, delta smelt, and largemouth bass as well as different categories of zooplankton, phytoplankton, and detritus. The level of resolution of such models depends on the availability of data. It would be possible to use such a model to trace the indirect dependencies of harvest in the bay-delta ecosystem, although Bauer did not report such results.
Models such as Bauer’s (2010), however, cannot account for evolutionary and behavioral changes in response to stressors. For example, Pine et al. (2009) report several examples of fishery ecosystems that exhibited fundamentally different responses to management perturbations than those predicted by ecosystem models. In two examples cited by Pine and colleagues, managers took action to increase the prey abundance for piscivorous fish in an effort to increase harvests of the piscivorous fish to anglers. However, increased prey abundance actually led to decreased abundance of the desired predators because of competition between the juvenile stages of the predator and the prey for a shared zooplankton response. The central message in these studies is that ecosystems are highly nonlinear systems that are sensitive to multiple inputs such that simple dependencies of A gives B gives C may fail.
Conclusion
Exploitation is a significant structuring agent in a diverse range of ecosystems from small freshwater ponds to ocean basins. However, we lack definitive evidence that exploitation is a major stressor in the San Francisco Bay-Delta system, especially since the harvest of Chinook salmon and steelhead in California has been tightly controlled recently (green sturgeon, as a listed endangered species, may not be taken). That is not to say that
exploitation could not be a principal stressor, but simply that we lack the empirical collection programs and integrated modeling programs to determine the importance of exploitation in this system.
Hatchery-Related Effects on Anadromous Salmonids in the Central Valley
In the 2009 draft Recovery Plan for Central Valley Chinook salmon and steelhead, NMFS (2009b) identified hatchery effects as a stressor contributing to, or implicated in, the decline of ESA-listed anadromous salmonid species in the Central Valley. Even so, they acknowledge the positive roles that hatcheries have provided in certain circumstances. The current status of wild populations is a critical consideration when assessing benefits and risks associated with hatchery production. For the three ESA-listed salmonids, population structure today is greatly diminished compared to their historical status. According to NMFS (2009b), historically the Sacramento River winter-run Chinook salmon evolutionary significant unit (ESU18) was composed of four populations. It now consists of a single population, which depends on hatchery production. The Central Valley spring-run Chinook salmon ESU had as many as 18 or 19 total populations. Today there are three. NMFS (2009b) hypothesized that historically 81 independent populations of steelhead were dispersed throughout the region; today there are perhaps 26 populations.
Winter-Run Chinook Salmon
Given the depressed state of this ESU, NMFS recognizes the need to rely on the Livingston Stone National Fish Hatchery (LSNFH) near Red Bluff, California, as part of the overall conservation strategy. The population declined from nearly 100,000 returning adults per year in the late 1960s to fewer than 200 in the early 1990s (Good et al. 2005). In the past two decades the number of returning adults have ranged from 2,542 in 2007 to a high of 17,153 in 2006 (NMFS 2009b).
NMFS considers LSNFH to be a good example of a conservation hatchery whose intent is to increase genetic diversity and minimize domestication of the hatchery progeny. Even so, Lindley et al. (2007) identify hatchery influence on the wild population as a potential concern with regard to genetic diversity. Even a small contribution of hatchery fish to the natural spawning
![]()
18 NMFS uses the term “ESU” as a basis for identifying a “distinct population segment” as specified in the Endangered Species Act, under which a DPS of vertebrates is included in the definition of “species for the purposes of the act.” The Endangered Species Act does not define a “distinct population segment,” but both it and the ESU are smaller evolutionary units than species or subspecies.
population could compromise the long-term viability of the population and increase the probability of extinction. Furthermore, Lindley et al. (2007) concluded that the winter-run population “is at moderate extinction risk according to the PVA [population viability analysis],” and at low risk according to other criteria (i.e., population size, population decline, and the risk of wide ranging environmental catastrophe).
Although in the recovery plan NMFS did not explicitly indentify hatchery effects as a key stressor for this species, it expressed concerns as the proportion of hatchery fish on spawning grounds increases. Lindley et al. (2007) reported, based on unpublished data from NMFS, that since 2001, the hatchery-origin winter-run Chinook salmon from LSNFH “made up more than 5 percent of the natural spawning run,” and in 2005 it was more than 18 percent. As the percent hatchery contribution on the spawning grounds rise, so will concerns regarding the potential for negative genetic impacts, given there is only one wild population unit remaining.
Spring Chinook Salmon
NMFS estimates that historically there were up to 600,000 wild spring Chinook adults returning to the Central Valley and its tributaries. Since 1970, the number of hatchery and wild fish returning has generally ranged from 3,000 to 30,000 individuals each year. Hatchery fish are a substantial component of this run. The Feather River Fish Hatchery (FRFH), the only spring Chinook hatchery, was established in 1967. The target production is 2 million smolts released annually.
The release strategy at FRFH appears to have promoted excessive straying (that is, adults returning to a different stream from where they hatched). Up to 1 million smolts have been regularly released in San Pablo Bay, increasing the probability of returning fish straying throughout the Central Valley. In fact there is direct evidence of pronounced straying of spring-run Chinook salmon. The NMFS Recovery Plan (NMFS 2009b) reported that up to 20 percent of the sport catch in the American River are of FRFH origin. Cramer and Demko (1997) estimated that half of the hatchery-reared spring-run Chinook salmon returning to the Feather River did not return to the hatchery but spawned naturally in the river. The committee found no information on the spawning success and productivity of that population component.
Whether these observed distributions reflect true straying or wandering, i.e., detouring but eventually ending up in the natal stream, has not been determined. Straying has more negative implications since fish do not return to their natal streams. A wandering fish could eventually return to its natal stream following attraction to localized favorable water conditions, unless of course it is intercepted in a fishery. Furthermore, in the
past, outdated hatchery practices likely promoted the unintended mixing of two Chinook salmon races at the hatchery, resulting in hybridization (CDWR and PSMFC 2004). At times, both populations were together in the hatchery complex due to periodic temporal overlap of returning spring and fall-run populations at the hatchery. Current practices strive to minimize such mixing.
Central Valley Steelhead
For steelhead, the increase in Central Valley hatchery production has reversed the wild-to-hatchery ratio of the steelhead population since the 1950s. McEwan (2001) quoted Hallock et al. (1961) as estimating that historically, 1 to 2 million adult steelhead returned to the Central Valley. In the 1960s those numbers dropped to near 40,000 (CDFG 1996). And by 1996, fewer than 10,000 returned to the system. In the 1950s, 88 percent of the population was composed of naturally produced fish (McEwan 2001), decreasing to an estimated 23 to 37 percent naturally produced19 fish in recent times (Nobriga and Cadrett 2003).
The NMFS recovery plan explicitly identifies hatchery effects as a major stressor contributing to the decline of Central Valley steelhead. It notes that hatcheries relied on nonlocal populations of steelhead in some of the hatchery programs. Early on, the Nimbus Hatchery on the American River imported fish from the Eel River, a coastal stream in northern California, and transferred that stock to hatcheries in the Central Valley. In the 1970s, the FRFH imported steelhead from Washington State and incorporated those into the breeding program. For these reasons NMFS judges the original gene pool has likely been compromised. Today such practices are generally avoided. One of the recommendations in the Joint Hatchery Review Report (CDFG and NMFS 2001) was to identify and designate new sources of steelhead brood stock to replace the current brood stock of Eel River origin.
Hatcheries—Benefit or Risk?
In the Central Valley, hatcheries have been established to offset the loss of wild production associated with dams that prevent access to 80 percent or more of the historical spawning and rearing habitat. The role of hatcheries in fisheries management has been a continuing topic of debate,
![]()
19 Naturally produced fish are the progeny of fish that spawned in the wild, whatever the origin of the parents.
particularly regarding anadromous salmonids. Do perceived benefits outweigh risks to wild populations?20
The extent to which hatcheries or hatchery fish have contributed and will continue to contribute to the decline of wild populations is difficult, if not impossible, to ascertain. There are often-cited benefits and risks associated with reliance on hatchery programs to satisfy fishery demands and supplement production of depressed wild populations. Devising strategies to achieve an acceptable balance among the risks and benefits is a constant source of debate. Waples (1991) stressed hatchery risks and argued that releasing large numbers of hatchery fish could adversely affect wild Chinook salmon and steelhead through various mechanisms such as hybridization, competition between hatchery and wild fish for food and other resources, predation by hatchery fish on juvenile wild fish, and the effects on wild fish of increased fishing pressure as a result of increased hatchery production.
Brannon et al. (2004) concluded that hatchery fish have an important role in recovery and supplementation of wild stocks. Riley et al. (2004) reported that small-scale releases of hatchery-reared smolts of Chinook or coho (Oncorhynchus kisutch) salmon had few significant ecological effects (density, group size, and microhabitat use) on wild salmonid fry in small, coastal Washington streams, particularly when the densities of wild salmonids are relatively low. They acknowledged the numbers of fish released was considerably smaller than most hatcheries release. More than two decades ago Hillman and Mullan (1989) observed that the release of numerous hatchery fish was associated with a decrease in abundance of wild salmonids in the Wenatchee River, Washington.
More recently, Araki et al. (2008) and Christie et al. (2012) reviewed and analyzed information on genetic effects on salmonids of hatchery rearing. They concluded that domestication selection can produce significant reductions in fitness in steelhead, Atlantic salmon (Salmo salar), and coho salmon. The declines can occur surprisingly rapidly, in as little as one or two generations, even in hatchery stocks derived only locally, that is, derived from the stream into which their progeny will be released. The National Research Council (2004) concluded that even with the best possible hatchery practice, domestication selection cannot be entirely eliminated. In addition, it is almost impossible to avoid selection for changes in run timing, especially in the diversity of run timing within populations (NRC 2004).
![]()
20 “Wild” fish often are defined as being the second generation of naturally produced fish (e.g., NRC 2004, McElhany et al. 2000). It is difficult or impossible to identify pristine populations of anadromous salmonids in the continental United States, that is, populations that have never been altered by introduction of genes from hatchery fish or fish from other populations. Such populations likely are rare, if they exist at all (NRC 2004).
From an ecological perspective, Mobrand et al. (2005) concluded that hatcheries must be considered part of the ecosystem in terms of biomass input, effluent, and predation-competition dynamics involving wild fish. Ecological and genetic interactions involving wild and hatchery anadromous salmonids are of concern in terms of competition for habitat and resources, predation on smaller life stages, interbreeding, and reproductive success.
Fishery managers regularly confront the dilemma of satisfying commercial and sport fisheries with abundant hatchery production, while simultaneously attempting to conserve threatened and endangered wild populations. An important consideration is the often unintended consequence of harvesting depressed wild stocks in a mixed-stock fishery fueled by abundant hatchery fish. Such a “mixed-stock” fishery tends to further depress less-productive stocks at the expense of the more-productive, often hatchery-based, stocks. This matter was discussed in detail by the NRC in an earlier report (NRC 1996). Since the goals of producing large numbers of fish for exploitation and conserving the genetic variability of wild populations conflict, and because relatively little harvest of steelhead and salmon in California is currently permitted, decisions will need to be made about the purposes of the national fish hatcheries. This, indeed, echoes a recommendation of the Hatchery Scientific Review Group (2009) (see below).
More than a decade ago, Congress recognized the inherent conflict between boosting hatchery production to supply fisheries and the obligation to protect and conserve depressed wild stocks, and thereby funded a hatchery reform project in Washington State in the year 2000. The ensuing Hatchery Scientific Review Group (HSRG) formulated guidelines for balancing the needs of both hatchery-produced and wild salmonids in the Columbia River system.21 In particular, the HSRG concluded that “[h]atchery fish cannot replace lost habitat or the natural populations that rely on that habitat. Therefore, hatchery programs must not be viewed as surrogates or replacements for lost habitat, but as tools that can be managed as part of a coordinated strategy to meet watershed or regional resource goals” (HSRG 2009).
But even in light of these numerous concerns regarding risks and negative effects associated with hatcheries, NMFS (2009a) sees the need to continue reliance on them in the broader recovery strategy and recommends that a hatchery supplementation plan be formulated. The Biological Opinion (NMFS 2009a) notes (LF 2.2),
In consultation with the NMFS Southwest Fishery Science Center, Reclamation shall develop and implement a long-term population supplementa
![]()
21 The HSRG’s reports are at http://hatcheryreform.us/hrp/welcome_show.action. (The HSRG began a review of the Klamath River and Central Valley systems in 2010; reports were not yet available in early 2012.)
tion plan for each species and fish passage location identified in V. Fish Passage Program, with adult recruitment and collection criteria developed with consideration for source population location, genetic and life history diversity, abundance and production…. The plan shall identify wild and/or hatchery sources for adult reintroductions and long-term supplementation, and the specific NMFS-approved hatchery management practices that qualify a hatchery for conservation purposes. Species-specific conservation hatchery programs may be developed to supplement reintroductions and maintain long-term performance standards for abundance and viability.
Conclusion
The committee recognizes the risks that have been imposed on wild salmonid populations by hatchery programs to date, and shares the concerns voiced by the scientific community and in the NMFS recovery plan. However, because negative effects of hatcheries are difficult to observe, the committee cannot reach a conclusion as to whether and how much hatcheries have contributed to the decline in wild populations in the Central Valley. In fact, the NMFS recovery plan asserts that for winter-run Chinook the LSNFH is one of the most important reasons the winter run persists. Importantly, the committee sees the need to follow recommendations of the NMFS Recovery Plan to formulate a new comprehensive hatchery program, and adopt the Viable Salmonid Population guidelines (McElhany et al. 2000) as guiding principles for long-term recovery. The committee expects continued reliance on hatcheries in the future, given the limited amount of productive spawning and rearing habitat that will likely be available.
Araki et al. (2008) concluded that the “general finding of low relative fitness of hatchery fish, combined with studies that have found broad scale negative associations between the presence of hatchery fish and wild population performance (e.g., Hoekstra et al. 2007), should give fishery managers pause as they consider whether to include hatchery production in their conservation toolbox.” The NRC (2004) concluded that despite a more than 130-year history of stocking,
[t]he available information is not sufficient to conclude whether hatcheries in Maine can actually help to rehabilitate [Atlantic] salmon populations, whether they might even be harming them, or whether other factors are affecting salmon so strongly that they overwhelm any good that hatcheries might do.
How do those conclusions apply to the steelhead and Chinook salmon of the Central Valley? Atlantic salmon in Maine, for example, have had fewer than 2,000 adult returns in recent years, including hatchery fish, at least an order of magnitude fewer than the number of returning Chinook
salmon in the Central Valley, but more comparable to the number of steelhead returns. In addition, steelhead, Atlantic salmon, and coho salmon typically spend a year or more as juveniles in freshwater before migrating to the ocean, whereas Central Valley Chinook juveniles typically spend less than a year. The committee cautions against applying results from one species to another without careful consideration of potential differences between them, and reiterates the difficulty of confidently ascribing observed changes in salmonid populations to hatchery effects. Nonetheless, the committee concludes that the cautionary notes developed from studies of Atlantic salmon in Maine and coho and steelhead in the western United States are generally applicable to all of the anadromous Central Valley salmonids; for Chinook, especially, more specific information would be valuable. The committee judges that adoption of HSRG guidelines under a unified hatchery management plan will reduce (but not eliminate) risk to wild populations from hatcheries, and probably represents the most viable option for maintaining populations of salmonids in the Central Valley unless or until other methods are found to increase the productivity of wild populations.
Ocean Conditions
Ocean conditions have a significant impact on all fish that pass through the delta and reside in the ocean during part of their life cycle. Particularly affected are salmon, steelhead, and sturgeon, which are anadromous. Their adult stages occur in saltwater, they pass through the delta to spawn in streams, and the juveniles pass through the delta on their migration to the ocean. The major mechanism by which the ocean affects anadromous fish, and in particular salmon, is known as bottom-up forcing, in which patterns in atmospheric temperature, wind, and precipitation drive ocean temperatures, mixing, and currents, which in turn control growth and advection of plankton that provide food for salmon (Batchelder and Kashiwai 2007).
Year-to-year variability in coastal conditions affects early ocean survival of delta salmon (Williams 2006b, Williams in press) and variability in ocean indices appears to be increasing (N. Mantua, U. Washington, unpublished data cited in Lindley et al. 2009). Such variability appears to have contributed to a collapse of the 2004 and 2005 brood years of Central Valley fall Chinook. When in 2005 and 2006 the broods passed though San Francisco Bay and into the Gulf of the Farrallones, conditions were poor. The juveniles experienced periods of weak upwelling, warm sea surface temperature, and low density of prey (Lindley et al. 2009). The estimated survival from hatchery release to age 2 was only 3 percent of the survival of the 2000 brood. Lindley et al. (2009) proposed that the impacts of year-to-year variations in coastal conditions are amplified because releases of juvenile hatchery fall Chinook salmon are correlated among nearby hatcheries.
Thus, the combined effects of hatchery-synchronized juvenile outmigration and a possible increase in ocean variability may lead to more booms and busts in the fisheries.
The longer-term effects of bottom-up forcing are more difficult to assess, but this is an active area of research under the coordination of the North Pacific Marine Science Organization (PICES 2010). The problem is being considered across several temporal and spatial scales. The Pacific Decadal Oscillation (PDOs), which characterizes the decadal scale variations in ocean temperature and currents, has a significant impact on the coastal habitat of west coast salmon. During the warm phase of the PDO, there is less advection of cold-water zooplankton species from the north and strong advection of warmer-water species from the west (DiLorenzo and Minobe 2010). The cold-water species have higher lipid content and are thus more nutritious for salmon, which is thought to improve early ocean survival of the salmon (Peterson and Tadokoro 2010). PICES studies also are focusing on the effect of large-scale climate variability on the lower trophic levels (see also Cloern et al. 2010). While the PDO is correlated with many west coast salmon stocks, a clear correlation with Central Valley Chinook salmon has not been found (Botsford and Lawrence 2002). The lack of correlation might involve unique oceanic conditions in the Gulf of the Farallones (Williams 2006b). San Francisco Bay and the Gulf of the Farallones lie at the southern boundary of the ocean habitat of salmon. Furthermore, the boundary is predicted to shift north with climate change (Irvine 2010).
In any case, studies to identify the effects of ocean changes on fisheries are in their initial stages (DiLorenzo et al. 2010). Studies are identifying detailed mechanisms that relate past changes in ecosystems to climate forcing (Lluch-Coat et al. 2010). While they are focused on the ocean boundary ecosystem, there is no emphasis on the southern boundary where Central Valley salmon and steelhead first enter the ocean. Furthermore, little information is available on climate impacts on the high seas habitat of steelhead and some runs of Chinook. The effects of climate warming and CO2 on ocean chemistry and the resulting effects on marine life are of concern, but it is too early to draw conclusions as to the likely responses of ecosystems (Denman et al. 2010).
In conclusion, despite wide-ranging and international research on the effects of changes in ocean conditions on fish and fisheries, there is little focus on the Central Valley stocks. This is unfortunate because these stocks enter the ocean at the southern boundary of the habitat and so it is plausible that ocean changes will have a significant impact on them.
Disease
Fish are constantly exposed to bacterial, fungal, protozoan, and viral pathogens but are generally protected from disease by a series of defense systems. The first line of defenses are the skin, scales, and mucus layers that trap and inhibit growth of pathogens. Pathogens that breach these systems are attacked by specific and nonspecific immune systems (Iwama and Nakanishi 1996). The ability of pathogens to overcome these defense systems and cause disease depends on abiotic, biotic, and genetic factors (Snieszko 1973). Disease may be enzootic, persisting in the population without significant impacts, or occur as short-term epizootic disease, which may have a significant impact on a population. Furthermore, fish exposure to pollutants and contaminants can lead to immunosuppression and increased susceptibility to infection (Arkoosh et al. 1998). Disease spread within a population also depends on the proximity of noninfected to infected individuals, for example, as occurs in hatcheries and at passage facilities where fish densities are high. Disease is also spread to offspring through inbreeding of infected and noninfected fish (NMFS 2009a). Analyses of threats to delta fish species typically mention disease as a cofactor with biotic and abiotic stress (e.g., NMFS 2009a, Baxter et al. 2008). In these situations, disease may occur as a result of reduced immunocompetence (i.e., the ability to ward off disease).
Generally, immunocompetence is lowest in young and old fish. Additionally, immunocompetence decreases during periods of hormonal stress, e.g., parr-smolt transformation of salmon and sexual maturation (Tatner 1996). Xenobiotic stressors such as metals, aromatic hydrocarbons, and pesticides reduce immunocompetence (see discussion of contaminants). Elevated temperature can have a major effect on stress response (Schreck 1996), which affects immunocompetence, and NMFS (2009a) notes that elevated temperature associated with climate warming may lead to increased disease in salmon.
In spite of the great potential impact of disease on delta fish populations, evidence for significant direct impacts of disease, or the impacts associated with containments, is mixed. Whirling disease caused by the parasite Myxobolus cerebralis is established in California salmonid populations but has been in decline and epizootic infections have only been reported in hatchery populations (Modin 1998). Infectious hematopoietic necrosis virus (IHNV) is common in juvenile hatchery salmonids but horizontal transmission of the Sacramento River strain of IHNV to wild cohorts appears to be a low ecological risk (Foott et al. 2006). High temperatures and fish densities induced an outbreak of disease in Chinook salmon oversummering in Butte Creek and resulted in prespawning mortalities between 20 and 60 percent (NMFS 2009a). However, in general wild salmon tend to
be less susceptible to disease than hatchery salmon (NMFS 2009a). Histopathological and viral evaluation of young longfin smelt and threadfin shad indicated no histological abnormalities associated with toxic exposure or disease (Foott et al. 2008). Adult delta smelt collected from the delta exhibited little histopathological evidence for starvation or disease, while there was some evidence of endocrine disruption (Teh et al., unpublished reference called out but not referenced by Baxter et al. 2008). However, studies in Suisun Bay reported fungal infection in yellowfin goby and viral infections in inland silverside and juvenile delta smelt. High occurrence of parasitic infection and inflammation and muscle degeneration were reported for striped bass (Baxter et al. 2008). Evidence suggests these infections may have been associated with the transfer of xenobiotics on larval striped bass in the San Francisco Estuary (Ostrach et al. 2008). Irrespective of these documented instances of pathogens, little information exists to quantify changes in infection and disease-associated mortality effect in Central Valley salmon (NMFS 2009a) and other species.
In short, the studies to date do not suggest that disease by itself or associated with contaminants has a major impact on the population levels of anadromous and estuarine fish that migrate through or inhabit the delta.
Multiple Stressors and the Pelagic Organism Decline
The pelagic organism decline (POD) was the simultaneous decline beginning in 2002 of the abundance indices of delta smelt, longfin smelt, threadfin shad, and juvenile striped bass. The POD study is a major effort at determining the role of the different stressors in causing the fish declines. A POD management team was established in 2005 by the IEP. Roughly every 2 years, the POD management team synthesizes the results of the various research projects to push toward answering the overarching question of what caused the POD (IEP 2006, Baxter et al. 2008, 2010); a final report is due in 2012-2013. In the periodic reports from the POD management team, the POD results were described by stressor (driver) for each life stage and season for each of the four species.
Initially, a single conceptual model was proposed, which was followed by refinement as species-specific models. The conceptual models were organized as life-cycle diagrams, with the definition of life stages and time periods related to the commonly used monitoring data (e.g., fall midwater trawl, summer tow net survey, 20-mm survey). The list of drivers (stressors) has increased during the POD study, paralleling the increasing complexity in the evolving conceptual models. Initially, the POD synthesis reports used a generic life-cycle conceptual model with three drivers (toxic substances, exotic species, and water projects). This was expanded by the IEP’s POD Management Team “to species-specific models and nine driv-
ers: (1) mismatch in time and space of larvae and their key prey items; (2) reduced habitat area and volume; (3) adverse water movement/transport; (4) entrainment; (5) toxic effects on fish; (6) toxic effects on fish food items; (7) harmful M. aeruginosa blooms; (8) C. amurensis effects on food availability; and (9) disease and parasites.”
Quickly, it became apparent that a “smoking gun” (i.e., a single driver that was the cause) was not present and the philosophy shifted more toward evaluating the effects of multiple stressors acting together. Two statistical analyses (Mac Nally et al. 2010, Thomson et al. 2010), done partly with funding coordinated by the IEP, concluded that important covariates correlated to the fish abundance indices prior to the POD were no longer related to the abundance indices observed after the POD. Using the results of various studies, the overall POD synthesis efforts by the management team identified some potential drivers that were not strongly related to abundances, which helps to constrain the problem, and illustrates the difficulties in relating drivers to population dynamics of the fish species.
Most recently, the view from the POD management team has evolved with more attention paid to the longer-term declines and the notion of an ecological regime shift superimposed on the effects of the multiple stressors. A regime shift is a relatively sudden, large-scale change in the state of ecosystem from one stable configuration to another due to nonlinear responses to slow changes in drivers (Andersen et al. 2009). The POD is a recent decline within long-term declines for each of delta smelt, juvenile striped bass, and longfin smelt.
The POD study is an excellent example of the type of synthesis that is needed to examine the effects of multiple stressors on fish species declines. The POD study involved many people and used strategic planning with conceptual models to design the study elements and then piece the results together. A logical next step to the POD study is to further evolve from the purely qualitative, conceptual formulations of species life cycles to more-quantitative life-cycle modeling analyses.
The POD effort has contributed to a major shift in the thinking of stressor effects on fish species in the delta. The long-held earlier idea that a single stressor (e.g., entrainment) must be the cause has changed to now examining the simultaneous, and potentially interactive, effects of multiple stressors. This is a landmark change in thinking. Whether the latest idea of an ecological regime shift has a similar impact on the scientific thinking about the declines in fish in the delta community remains to be determined. Some stressors can be eliminated, but the remaining stressors are difficult to rank because their occurrence overlaps and their effects can be nonlinear, episodic, and interactive.
Priority Stressors for ESA-Listed Species
The committee was asked to attempt ranking the importance of various stressors with regard to their importance in affecting survival, productivity, and ultimately recovery of endangered fish species. The following is a discussion and evaluation of stressors for listed species.
Anadromous Salmonids
At least two forums have attempted to characterize and score the importance of various stressors on ESA-listed anadromous salmonids in the Central Valley; the NMFS Draft Recovery Plan and the Delta Conceptual Models22 for those species. The NMFS approach is very detailed and specific as to species and population unit, life stage, and river locale. Although very thorough and extensive in their treatment, the committee encountered no concise distillation by broader stressor categories. Since the charge of this committee is coarser and broader, the committee could not readily distill their evaluations to align well with our approach to Central Valley stressor impacts.
The Delta Conceptual Models considered stressor impacts at a higher level, more consistent with our charge, and we looked to those for reference, against which to judge our assessments of stressors. However, they had some limitations for our purposes. The stressor scoring focused on ten categories of stressors, for each of four salmon life stages. Each was scored from low (1) to high (4) in terms of three parameters: understanding, importance, and predictability. Although a promising template, many of the cells in the matrix were not scored, leaving the assessment still open ended. Furthermore, many of the environmental and anthropogenic factors considered candidate stressors in this report were not treated in that document. Thus, the committee concludes that in their present form the salmonid conceptual models provided a still incomplete picture with regard to assessing the importance of a broad spectrum of stressors.
The committee treats the anadromous salmonids as a species complex in the ranking discussions. The loss of access to over 80 percent of the historical habitat has led to greatly reduced carrying capacity and simplified population structure. These outcomes limit abundance, productivity and resiliency, even if a variety of other stressors are relaxed.
Unless these fundamental constraints are relaxed, recovery goals will be very difficult to attain and the populations might even be in danger of extinction, especially in the face of expected climate change. Altering pump operations or providing an alternative water conveyance system will do
![]()
22 See http://www.science.calwater.ca.gov/drerip/drerip_index.html. Accessed July 17, 2012.
little to offset the dramatic effects of habitat loss and deficiencies in existing population structure. The successful reintroduction of salmonids to select, expansive, and productive watersheds will provide needed increased carrying capacity, provide access to thermally acceptable areas, and enable the selection for and expression of new life history patterns and accompanying establishment of new population units. NMFS recognizes this as reflected in the draft Recovery Plan and has identified candidate watersheds that are currently inaccessible to salmonids. Feasibility studies for reintroduction remain to be formulated. The strategy of reintroducing anadromous salmonids to drainages upstream from dams lacking fish ladders is being implemented in the Pacific Northwest (PNW), with notable success. Sockeye and coho were successfully reintroduced above two dams on the Baker River in Washington State. On the Lewis River, efforts are under way to reestablish naturally producing salmon populations upstream of Swift Dam. The same type of effort has been ongoing on the Cowlitz River for over a decade. Numerous other sites in the PNW are now being assessed as to feasibility (AECOM and BioAnalysts 2010). The implementation involves collecting adults at the base of the dam and transporting them above the barrier. But the most technically difficult task involves the design of a collection system that can safely and effectively intercept and route smolts around the structure (AECOM and BioAnalysts 2010).
Dam removal can be an effective strategy for increasing the area of habitat for diadromous fishes, but it is not simple or inexpensive, especially for large dams (Heinz Center 2002, NRC 1996, 2004). In a water-short region like the delta watershed, it seems unlikely that any large dams will be removed soon. In addition, large dams can be used to mitigate the adverse effects of increasing temperatures in the waters below them by providing for the release of cold water. Removal of small dams high in the watershed might provide some benefit in combination with other strategies, but the feasibility and desirability of doing so would need to be weighed carefully against the costs and other disadvantages (NRC 2004).
Green Sturgeon
Green sturgeon (Acipenser medirostris) is a diadromous species that occupies different Central Valley habitats depending on life stage and season (Figure 3-10). The Central Valley subpopulation is very small. Moyle (2002) estimated that between 140 and 1,600 adults occupy the system each year, while Israel and May (2010), using molecular kinship analysis of various life stages in the upper Sacramento River, estimated that their results could be accounted for by as few as 10 to 28 spawning adults above the Red Bluff Diversion Dam each year. The Central Valley subpopulation
FIGURE 3-10 Schematic of the life cycle of green sturgeon.
SOURCE: Beamesderfer et al. (2007).
is part of a southern population segment that is genetically distinct from a northern population segment (Israel et al. 2009).
Adult green sturgeon (13-16+ years old) migrate into the bay-delta system in winter months. Most adult sturgeon undertake spawning migrations up the Sacramento River beginning in March and reach upriver locations (adjacent to and above Red Bluff Diversion Dam) in late spring and early summer (Beamesderfer et al. 2007, Israel and Klimley 2008, Heublein et al. 2009).
It is believed that sturgeon spawn in this area in stretches of the river with high flow over bedrock. Fertilized eggs hatch in 6-8 days, and then larvae initiate feeding and disperse downstream in a nocturnal diel migration. Following metamorphosis to juveniles, sturgeons adopt a benthic orientation, feeding on invertebrates while they move slowly downstream to lowland and estuarine habitats. By 3 years of age, juveniles move into the coastal ocean (Beamesderfer et al. 2007), where they migrate widely, co-occurring with the northern population segment in the estuaries of the Pacific coast of North America (Lindley et al. 2008).
Green sturgeon populations are susceptible to stressors in several life stages. Israel and Klimley (2008) and the NMFS Biological Opinion (NMFS
2009a) identified critical factors in the larval early life period in the Sacramento River habitat: (1) warm water temperatures, (2) insufficient flows, (3) decreased dissolved oxygen, (4) lack of rearing habitat, and (5) increased predation. Although such studies rarely address the issue, bioaccumulative contaminants like selenium and mercury also pose a particular risk for this species. The risks stem from the sturgeon’s high trophic position in the benthic food web, the importance of life stages most at risk from these reproductive toxicants, and the poor demographic compensational abilities of this long-lived, slowly reproducing species. Because the southern distinct population segment is listed under ESA, recovery action may extend beyond the San Francisco Bay Delta. Using elasticity analysis, Heppell (2007) identified sensitive life history stages and concluded that reductions in bycatch would be the most effective way of restoring green sturgeon. Mora et al. (2009) developed a green sturgeon habitat suitability model and concluded that broad reaches of the Sacramento–San Joaquin system now are unavailable because dams block formerly suitable spawning habitat. However, they caution that their conclusions are fraught with uncertainties because of the complex impact of dams on the ecology and hydrology of impounded rivers. In addition, they comment that efforts to regulate flow to benefit endangered salmonids may have had negative effects on sturgeon.
Overall, the committee makes the following points from a review of the green sturgeon literature:
1. Recent genetic and hydroacoustic tagging evidence continues to support the existence of two distinct population segments: a southern DPS that spawns in the bay delta and in particular in the Sacramento River, and a northern DPS that spawns in rivers in Oregon and Washington. However, these same data also clearly show that the two DPSs mix throughout the range at all times except while spawning. This suggests that restoration of the southern DPS in the bay delta must also keep in mind the status of green sturgeon throughout its range.
2. The value of subadult sturgeon to future population growth is substantial and thus efforts to reduce the bycatch and other incidental sources of mortality on this life history stage should continue. There has already been a concerted effort that has reduced bycatch from thousands of fish in the second half of the 20th century to hundreds of fish today.
3. The temporal and spatial distribution of sturgeon in the Sacramento–San Joaquin river system is relatively well described and should inform management efforts to maximize the suitable habitat. However, there are inherent trade-offs in trying to promote maximum habitat quality for different species of concern. What may be good
for salmon may not be beneficial to sturgeon or delta smelt. These trade-offs should be carefully identified and considered when making management decisions for each species.
4. Our knowledge of the distribution and habitat use of larval and juvenile sturgeon is particularly weak. Knowledge would be advanced by development of the kind of geospatial habitat quality model produced by Mora et al. (2009) for adult green sturgeon. This approach uses information for sturgeon throughout their range to inform decision for the bay delta.
5. The vulnerability of both green and white sturgeon to bioaccumulative contaminants like selenium and mercury is well known (see contaminants section). While habitat is clearly crucial in determining the fate of this species, recovery could be slowed by the existing levels of selenium and mercury contamination in the bay delta. Exacerbation of selenium or mercury contamination would increase risks to this species in particular, and with some possibility of the extirpation of the relict populations that currently inhabit the bay.
Delta Smelt
Several analyses have attempted to determine the importance of stressors affecting the population dynamics of delta smelt. While all the analyses generally agree on the list of potential stressors, the relative importance of the different stressors was highly variable across the analyses. These analyses are the DRERIP conceptual model, the POD synthesis study, three statistical analyses, and a life-cycle model. The analyses were not independent. They all used overlapping data sets, and the DRERIP conceptual model and POD synthesis study were qualitative and relied on the results of other analyses.
DRERIP
The Delta Regional Ecosystem Restoration Implementation Plan (DRERIP) was one of four regional plans intended to guide the implementation of the CALFED Ecosystem Restoration Element of CALFED.23 DRERIP developed a series of conceptual life-cycle models for key species, including delta smelt (Nobriga and Herbold 2009). Primary and secondary drivers were identified as factors affecting habitat, and primary and secondary stressors were identified that affect population abundance but operated through one or more drivers. Drivers determined to be affecting delta smelt abundance were sporadically high adult and larval entrainment, spring water temperatures reducing spawning season duration, warm summer-fall
![]()
23 See www.dfg.ca.gov/delta/erpdeltaplan. Accessed July 17, 2012.
water temperature, decreased summer-fall habitat area based on salinity and water clarity, and suppression of the food (zooplankton) base. A variety of stressors can contribute to each of these drivers. For example, water exports and Delta Cross Channel operations are stressors that affected the driver of entrainment, and introduction of the overbite clam is a stressor related to changes in the driver of food base.
Pelagic Organism Decline (POD)
The POD management team hypothesized that habitat degradation was the fundamental cause of the smelt decline through its effects on growth and reproduction (see Baxter et al. 2010 for details). Warm temperatures and low food quality and quantity during the summer slowed growth; smaller adults produced fewer eggs. Calanoid copepods have shown a long-term decline and the feeding area during the summer has been reduced due to warm temperatures and lower turbidity. Fall habitat, as measured by low salinity and high turbidity being good, has also showed a long-term decline; the specifics of how fall habitat relates to survival (via predation, disease, and food) are unclear. The shrunken fall habitat also places the smelt closer to the pumping facilities, which could increase entrainment in the following winter. High winter entrainment of adults during the POD years was superimposed on a low abundance comprising small individuals with low egg production. Entrainment of juveniles occurs during the spring and is generally higher during dry years (i.e., POD period), which overlapped with the years of high adult entrainment in winter. Blooms of toxic cyanobacteria and contaminants effects likely had less influence.
Statistical Analyses
Two analyses (Mac Nally et al. 2010, Thomson et al. 2010) used almost identical explanatory variables. Mac Nally et al. (2010) used a multispecies approach to examine whether the associations between the target species and other species and between target species and environmental covariates were consistent throughout the historical record. Thomson et al. (2010) focused on individual species and examined whether decreases in delta smelt, longfin smelt, juvenile striped bass, and threadfin shad abundance indices in the early 2000s were the continuation of longer-term trends (since the 1970s) or were more abrupt changes, and whether the covariates important before the changes continued to be important after the changes. The explanatory variables used for analyses of delta smelt included, among other variables, spring and summer biomass of calanoid copepods, chlorophyll a, spring X2, winter and spring exports, summer water temperature, PDO, biomass of Limnoithona copepodites and adults during the summer, water clarity, and mean catch rates of inland silverside and largemouth bass during the period July through September. Thomson et al. (2010) inferred that
a step change occurred in delta smelt and generally identified water clarity and winter volume of exported water as important, and that the importance of the covariates was not maintained after the step change. Mac Nally et al. documented biological and environmental associations involving delta smelt, but did not find that these associations changed after 2002.
Miller et al. (2012) reported the results of a third statistical analysis of delta smelt abundance indices and similar but not identical explanatory variables as Thomson et al. (2010) and Mac Nally et al. (2010). Miller et al. (2012) used the ratio of delta smelt abundance indices as a measure of survival (fall to summer, summer to fall, and fall to fall) and related these to covariates using a regression model that included density-dependence (i.e., a Ricker relationship) (Ricker 1954). Because of the potential for correlation among many of the covariates, Miller et al. (2012) used a complicated approach of subsetting the covariates and switching them to determine possible redundancies in detected relationships. They found associations between the fall-to-summer survival index and abundance indices in previous years, zooplankton densities, and entrainment. Summer-to-fall and fall-to-fall survival indices were related to previous abundance indices and zooplankton measures.
Life-Cycle Model
Maunder and Deriso (2011) developed a stage-based life-cycle simulation model of delta smelt population dynamics. They assumed possible density-dependent survival between life stages, and fit the model to the long-term monitoring data (summer tow net, fall midwater trawl). They then statistically searched for improved fit by allowing for explanatory variables (covariates) to be included as modifiers of stage survivals, singly and in combinations and either before or after density dependence. Using two of the fitted models, they then examined the importance of the explanatory variables, including entrainment. They concluded density-dependent survival was important for survival from juveniles to adults, and that covariates related to food abundance, water temperature, and predator abundance were most correlated with the interannual population variation. Adult entrainment also played a role in some of the final models selected.
Summary
These analyses used a variety of techniques applied to overlapping data sets, and they produced similar subsets of stressors as being correlated with the variability in delta smelt abundance indices. However, the different analyses did not converge with respect to the relative importance of individual stressors within the subset of generally important stressors.
Conclusions
While there are factors that all analyses agree are not important, there has been little agreement on the ranking among the potentially important factors for delta smelt. These analyses shared, to some extent, the same database but did not all use the same list of potential explanatory variables and sometimes used different numerical versions of the same explanation variable. The analyses identified various mixtures of important covariates, including spring and summer plankton biomasses, predator indices, water and spring exports, and fall water clarity.
Multiple stressors have affected and continue to affect the delta ecosystem and its biota, including the listed species of fishes. While some species have increased in the delta in the past few decades, listed fish species have continued to decline. The committee concludes that while it is possible to identify more-significant and less-significant stressors, a precise ranking of them, even for individual species, is not possible.
First, statistical evidence and models suggest that both flows and flow paths are critical to population abundance of many species in the bay delta. However, none of the statistical flow-abundance relations suggest the existence of thresholds (i.e., that if the position of X2 were to be allowed to remain far upstream for suitable [species-dependent] periods of time, there would be irreversible declines in fish populations or the near elimination of critical ecosystem processes like primary production of phytoplankton). However, it is clear that very dry periods can alter species composition in more permanent ways (Alpine and Cloern 1992, Jassby et al. 2002, Kimmerer et al. 2009).
Thus, it appears that if the goal is to sustain an ecosystem that resembles the one that appeared to be functional up to the 1986-1993 drought, exports of all types will need to be limited in dry years, to some fraction of unimpaired flows that remains to be determined. Setting this level, as well as flow constraints for wetter years, is well beyond the charge of this committee and accordingly we suggest that this is best done by the SWRCB, which is charged with protecting both water rights holders and the public trust.
The idea of developing operating policies based on statistical models highlights a fundamental challenge inherent to the substantial uncertainty of the observed relationships between flows, the position of X2, or any other abiotic factor. Design of most engineered systems tends to rely on optimization of performance, given some knowledge of the system, a practice that is known in seismic engineering as performance-based engineering. If critical aspects of flow and flow variability could be identified, for example,
a short period of time when a run of salmon moves through the delta or when a critical life stage of delta smelt is vulnerable to entrainment, the timing of pumping or the flow path of the diverted flow could be chosen so as to maximize the amount of water diverted while minimizing the effect of those diversions on at-risk species. Given the diverse set of organisms and processes that constitute the bay-delta ecosystem, the ultimate success of any approach targeted only to particular species seems doubtful. In contrast, broad ecosystem approaches, recognizing substantial uncertainty, are needed, although they might require more water. A risk of any approach is that long-term changes in the food web due to invasions or nutrient inputs or climate change might alter the influence of flow on the ecosystem; thus, continued monitoring is essential.
The hard decisions that will need to be made are ones of balancing different kinds of trade-offs. These will be matters of policy rather than being the result of a straightforward application of “good science.” Nonetheless, exactly because statistical correlations are not adequate to fully explain the responses of aquatic species to either flows or flow diversions (paths), continuing the effort to better understand the processes that control the implications of both flows and flow paths is essential into the future.
For migratory salmonids, and probably green sturgeon, significant stressors are the dams. They are impediments to passage, cause the loss of spawning and rearing habitat, change the abundance of predators, and affect temperature and flow. These effects limit abundance and productivity and reduce genetic diversity through extirpation of the vast majority of unique populations once present in the system.
Limited survival studies suggest migrating salmon and steelhead smolts incur substantial mortality during river and delta passage. Increasing passage of smolts through Yolo Bypass may be a viable action for Sacramento salmon populations. However, options for San Joaquin fish are less certain, because studies suggest that passage through the delta main channel and collection and transport at the pumps—the two main passage options—result in equally low survival.
Entrainment effects of SWP and CVP pumping are likely large in some years and, thus, act as an episodic stressor that has a significant adverse effect on delta smelt population dynamics, although it is very difficult to quantify the effects in simple ways. The flow path within the zone of influence of the pumps is especially important as a stressor for this species. Inflows to the bay are clearly important for longfin smelt. A series of papers on estimating entrainment impacts (Kimmerer 2008b, 2011; Miller 2011) provides a good example of careful and constructive critique and response that is desperately needed to further refine these important analyses. The dialog should continue, and the approach also is needed for other controversial analyses. The committee reemphasizes the need for life-cycle
modeling and a collaborative process to reduce the paralysis that can occur from the use of dueling models and to encourage cross-comparisons and cross-fertilization. The recent surge in life-cycle modeling for both delta smelt and salmonids is encouraging.
Changes in nutrient loads and concentrations in the delta and bay, especially those for nitrogen and phosphorus, are stressors of increasing concern from water-quality and food-web perspectives. Further simplification of this ecosystem is a serious concern if the impacts of such inputs increase because of failure to better remove nutrients from waste streams, climate change, or human-induced changes in flows. Toxic pollutants such as selenium also appear to be significant stressors, especially for sturgeon, with San Francisco Bay and the San Joaquin River being the areas of greatest concern. With appropriate investments both nutrients and selenium issues can be better managed, probably to the benefit of both functioning and structure in the delta and the bay. Examples of actions with a high likelihood of net benefits for the environment include the following:
1. A nutrient reduction plan that moves toward reducing all biologically available forms of nitrogen (especially) and phosphorus could benefit the delta and regions of the bay by addressing the increasing unwanted symptoms of eutrophication such as harmful algal blooms. However, as in any ecosystem effort, care will need to be taken to ensure that a change in nutrient ratios (e.g., N:P ratio) does not inadvertently favor other unwanted species, such as Brazilian waterweed.
2. A transparent plan for sustaining the effort to improve the in-valley solutions for the selenium issue is essential to ensuring that selenium inputs to the bay will not increase in the future.
3. Continued study is essential, including scenario building, of the ecological risks from water quality changes, especially selenium, of changing flow paths in ways that result in an increase of the ratio of San Joaquin River to Sacramento River water entering the bay.
The above stressors also interact with each other and with changes in salinity, turbidity, and freshwater discharge/flows resulting from hydrologic changes in the delta and its tributaries, changes that have been attributed to water exports, changes in land use, and changes in the morphology of the delta. The latter factor, caused by canalization and the abundance of hardened structures that also have eliminated tidal wetlands, has affected delta smelt by changing the aquatic habitats they occupy. But such physical changes in habitat occurred long before the most recent collapse of delta smelt and other pelagic organisms. Flow-related changes in habitat are more complex to understand and could have a more recent origin. There
is evidence that restoration of shallow-water physical habitat could be of value to the growth of POD species (e.g., if it imitates the Yolo Bypass). But care should be taken in assuming that such restoration can reverse the present decline in such species, given the lag between changes in the morphometry of the bay delta and declines in abundance. Again, support for better understanding the processes that link flows, habitat structure, and habitat characteristics such as salinity, turbidity, and temperature should remain a high priority.
Introduced species have caused dramatic changes in habitat, prey, and predators of the listed fish species in the delta. Determining the contribution of introduced species compared with the effects of other stressors to changes in the delta ecosystem is difficult, because some effects are local, multiple stressors vary simultaneously, and the data or models to directly link introduced species to observed ecosystem changes often are lacking. Introductions of nonnative species will continue because management controls that substantially reduce risk are difficult and expensive to implement. Changes in human activities and climate change could exacerbate the problem in the future. If solutions to problems caused by invasive species are possible, they will come from better understanding of the life cycles and vulnerabilities of these species. New technologies offer some possibilities for solutions as well (e.g., sterilization of ballast water with nitrogen gas; Tamburrri et al. 2002), as do adjustments in management designed to reverse the habitat characteristics that favor such species (e.g., varying salinities in the delta). However, all such proposals should be carefully evaluated for their feasibility. Early detection through monitoring is useful to prepare for likely changes to the ecosystem.
Largely because negative effects of hatcheries are difficult to observe, the committee cannot reach a conclusion as to whether and how much hatcheries have contributed to the decline in wild populations in the Central Valley. The committee judges that adoption of recent conservation guidelines under a unified hatchery management plan will reduce (but not eliminate) risk to wild populations from hatcheries, and probably represents the most viable option for maintaining populations of salmonids in the Central Valley unless or until other methods are found to increase the productivity of wild populations.
Coastal ocean productivity is one of the most significant factors determining the ocean survival of juvenile salmon and the number of adult salmon that return to spawn. Increased variability in coastal conditions expected with climate change may increase variability in Central Valley salmon and steelhead recruitment. When ocean conditions are unfavorable, the effects can be partially ameliorated by increasing the diversity of wild and hatchery salmon ocean entrance timing.
Currently, disease does not appear to be a significant stressor factor for
juvenile or adult salmon. However, with climate change and increasing river temperatures disease may be become a major factor in salmon mortality.
The real complexities added to the system by these factors, as well as the complexities added by interactions of all the above considerations, mean that ecological changes in response to engineering changes will not necessarily be linear. Policies should be based on more than just the presence or absence of linear, simple relationships between a stressor and its target(s). While we recognize that policies must move forward, a continuing, transparent effort to study, model, and track environmental changes and how they are influenced by those policies, is essential. Given the diversity of the challenges presented by “stressors” to the bay delta, better integration of the governance structures and science-policy dialogue, as suggested in other chapters of this report, is another important ingredient in addressing the stressor issues.
It is clear from consideration of the many stressors and their impacts that eliminating any one is unlikely to reverse declines in the listed species. Nor is it constructive when the advocates of the implications of one stressor use that advocacy to suggest their stressor is not important. Opportunities exist to mitigate or reverse the effects of many of the above stressors. To make it more likely that any actions to rehabilitate the ecosystem are cost effective, continued effects analyses, modeling, and monitoring will be needed.
Models will not eliminate controversy and they will not eliminate the need for information gathering in the field. Indeed, well-designed models should guide data collection. Nonetheless, modeling remains an essential part of scientific endeavors in the delta, especially as a way to test hypotheses and to improve understanding. Three-dimensional models that include salinity, temperature, and flow also would be helpful. While such modeling might not benefit decision making in the immediate future, it would help scientists to understand the complexities of the system better and provide a guide for conducting analyses and perhaps experiments to better understand complex interactions. Finally, hydrologic water-routing models for the entire system, covering the northern tributaries, the delta, and the demand areas in the south, should continue to be developed as a way to understand water budgets and long-term patterns of water use under various alternatives.
The CALSIM model is an example of such a model (e.g., Systech Water Resources, Inc. 2011). The CALSIM and other models were discussed in the NRC’s earlier report (NRC 2010). CALSIM, which has been developed over many years, is a good, system-wide water-routing model. However, it is a water-supply planning model. With climate change, it may need to be linked to watershed hydrology models to investigate the expected changes in hydrology. Besides a planning model, agencies may also need operational
models. These models should help improve operations and be able to route high water flows through the system under future conditions.
Multispecies trade-offs have been quantified in other restoration programs to find possible solutions to what appear to be irreconcilable conflicts. Quantifying trade-offs is a way to initiate discussions that cut across traditional barriers. It is essential in finding solutions and identifying next steps in addressing the challenges that face the bay-delta rehabilitation effort. The SERES24 report concludes with a lesson that is as applicable to the bay-delta as it is to the Everglades: “If the trade-offs inherent within the … system are not acknowledged, and management actions switch between the extremes of what is best for one group versus another, the outcome is likely to be more harmful than need be for all groups involved.”
Aasen, G. A. 2011. 2010 Fish Salvage at the Tracy Fish Collection Facility. California Department of Fish and Game, Bay Delta Region. Stockton, CA.
Adams, P. B., C. Grimes, J. E. Hightower, S. T. Lindley, M. L. Moser, and M. J. Parsley. 2007. Population status of North American green sturgeon, Acipenser medirostris. Environmental Biology of Fishes 79:339-356.
AECOM and BioAnalysts, Inc. 2010. Willamette Downstream Fish Passage Design Requirements Report. Final Submittal to Portland District Corps of Engineers. Portland, OR.
AFS (American Fisheries Society). 2004. Common and Scientific Names of Fishes from the United States, Canada, and Mexico. 6th Edition. AFS Special Publication 29. Bethesda, MD.
Alpine, A. E., and J. E. Cloern. 1992. Trophic interactions and direct physical effects control phytoplankton biomass and production in an estuary. Limnology and Oceanography 37:946-955.
Anderson, R., P. J. Cranford, C. W. McKindsey, P. Strain, B. T. Hargrave, W. K. W. Li, and W. G. Harrison. 2006. Cumulative and far-field fish habitat effects. Ottawa: Fisheries and Oceans Canada, Science, Canadian Science Advisory Secretariat.
Andersen, T., J. Castensen, E. Hernández-Garcia, and C. M. Duarte. 2009. Ecological thresholds and regime shifts: Approaches to identification. Trends in Ecology and Evolution 24:49-57.
Araki, H., B. A. Berejkian, M. J. Ford, and M. S. Blouin. 2008. Fitness of hatchery-reared salmonids in the wild. Evolutionary Applications 2008:342-355. Available at http://marking.fishsciences.net/_reports/Arakia_et_al_2008_Evol_App.pdf. Accessed June 4, 2012.
Arkoosh, M. R., E. Casillas, E. Clemons, A. N. Kagley, R. Olson, P. Reno, and J. E. Stein. 1998. Effect of pollution on fish diseases: Potential impacts on salmonid populations. Journal of Aquatic Animal Health 10(2):182-190. Available at http://www.science.calwater.ca.gov/publications/sbds.html. Accessed June 4, 2012.
![]()
24 See the food web section of the Everglades Restoration Synthesis of Everglades Research and Ecosystem Services (SERES); http://www.everglades-seres.org/Products_files/SERES_Food_Web_Review%20copy.pdf.
Arthur, J. F., and M. D. Ball. 1979. Factors influencing the entrapment of suspended material in the San Francisco Bay-Delta Estuary. Pp. 143-174 in San Francisco Bay: The Urbanized Estuary, edited by T. J. Conomos. San Francisco, CA: Pacific Division of the American Association for the Advancement of Science.
Augustin, L. N., J. L. Irish, and P. Lynett. 2009. Laboratory and numerical studies of wave damping by emergent and near-emergent wetland vegetation. Coastal Engineering 56:332-340.
Baskerville-Bridges, B., J. Lindberg, and S. Doroshov. 2002. The effect of light intensity, alga concentration, and prey density on the feeding behavior of delta smelt larvae. Pp. 219-228 in Early Life History of Fishes in the San Francisco Estuary and Watershed, edited by F. Freyer, L. Brown, R. Brown, and J. Orsi. Bethesda, MD: AFS.
Batchelder, H. P., and M. Kashiwai. 2007. Ecosystem modeling with NEMURO within the PICES Climate Change and Carrying Capacity program. Ecological Modelling 202(1-2):7-11.
Bauer, M. 2010. An ecosystem model of the Sacramento-San Joaquin Delta and Suisun Bay, California. Chico, CA: California State University.
Baum, J. K., and R. A. Myers. 2004. Shifting baselines and the decline of pelagic sharks in the Gulf of Mexico. Ecology Letters 7:135-145.
Baxter, R., R. Breuer, L. Brown, M. Chotkowski, F. Feyrer, M. Gingras, B. Herbold, A. Mueller-Solger, M. Nobriga, T. Sommer, and K. Souza. 2008. Pelagic organism decline progress report: 2007 synthesis of results. Interagency Ecological Program for the San Francisco Estuary, Technical Report 227. Available at http://www.science.calwater.ca.gov/pdf/workshops/POD/2007_IEP-POD_synthesis_report_031408.pdf. Accessed June 4, 2012.
Baxter, R., R. Breuer, L. Brown, L. Conroy, F. Feyrer, S. Fong, K. Gehrts, L. Grimaldo, B. Herbold, P. Hrodey, A. Mueller-Solger, T. Sommer, and K. Souza. 2010. 2010 Pelagic organism decline work plan and synthesis of results. Interagency Ecological Program for the San Francisco Estuary.
BDCP (Bay Delta Conservation Plan). 2007. Independent Science Advisors’ Report. Available at http://baydeltaconservationplan.com/Libraries/Dynamic_Document_Library/BDCP_ISA_Report_11-16-07.sflb.ashx. Accessed June 4, 2012.
Beamesderfer, R. C. P., M. L. Simpson, and G. J. Kopp. 2007. Use of life history information in a population model for Sacramento green sturgeon. Environmental Biology of Fishes 79:315-337.
Bennett, W. A., W. J. Kimmerer, and J. R. Burau. 2002. Plasticity in vertical migration by native and exotic estuarine fishes in a dynamic low-salinity zone. Limnology and Oceanography 47:1496-1507.
Boesch, D. F., E. Burreson, W. Dennison, E. Houde, M. Kemp, V. Kennedy, R. Newell, K. Paynter, R. Orth, and R. Ulanowicz. 2001. Factors in the decline of coastal ecosystems. Science 293:629-638.
Botsford, L. W., and C. A. Lawrence. 2002. Patterns of co-variability among California Current Chinook salmon, coho salmon, Dungeness crab, and physical oceanographic conditions. Progress in Oceanography 53(2-4):283-305.
Bouse, R. M., C. C. Fuller, S. N. Luoma, M. I. Hornberger, B. E. Jaffe, and R. E. Smith. 2010. Mercury-contaminated hydraulic mining debris in San Francisco Bay. San Francisco Estuary and Watershed Science 8(1):3. Available at http://escholarship.org/uc/jmie_sfews. Accessed June 4, 2012.
Brannon, E. L., D. F. Amend, M. A. Cronin, J. E. Lannan, S. LaPatra, W. J. McNeil, R. E. Noble, C. E. Smith, A. J. Talbot, G. A. Wedemeyer, and H. Westers. 2004. The controversy about salmon hatcheries. Fisheries 29(9):12-31.
Brooks, M. L., E. Fleishman, L. R. Brown, P. H. Lehman, I. Werner, N. Scholz, C. Mitchelmore, J. R. Lovvorn, M. L. Johnson, D. Schlenk, S. van Drunick, J. I. Drever, D. M. Stoms, A. E. Parker, and R. Dugdale. 2012. Life histories, salinity zones, and sublethal contributions of contaminants to pelagic fish declines illustrated with a case study of San Francisco Estuary, California, USA. Estuaries and Coasts 35(2):603-621.
Brown, C. L., F. Parchaso, J. K. Thompson, and S. N. Luoma. 2003. Assessing toxicant effects in a complex estuary: A case study of effects of silver on reproduction in the bivalve, Potamocorbula amurensis, in San Francisco Bay. International Journal of Human and Ecological Risk Assessment 9:96-119.
Brown, L. R., and J. T. May. 2006. Variation in spring nearshore resident fish species composition and life histories in the lower San Joaquin watershed and Delta. San Francisco Estuary and Watershed Science 4(1). Available at http://repositories.cdlib.org/jmie/sfews/vol4/iss2/art1. Accessed June 4, 2012.
Brown, L. R., and D. Michniuk. 2007. Littoral fish assemblages of the alien-dominated Sacramento–San Joaquin Delta, California 1980-1983 and 2001-2003. Estuaries and Coasts 30:186-200.
Brown, L. R., W. Kimmerer, and R. Brown. 2009. Managing water to protect fish: A review of California’s Environmental Water Account, 2001-2005. Environmental Management 43:357-368.
Burau, J., S. G. Monismith, and J. R. Koseff. 1993. Comparison of advective transport algorithms with an application to Suisun Bay. Pp. 1025-1030 in Hydraulic Engineering ‘93, edited by H. W. Shen, S. T. Su, and F. Wen. New York: American Society of Civil Engineers.
CDWR (California Department of Water Resources) and PSMFC (Pacific States Marine Fisheries Commission). 2004. Evaluation of the Feather River Hatchery Effects on Naturally Spawning Salmonids. Available at http://www.water.ca.gov/environmentalservices/docs/FR/newreports/effects_of_the_feather_river_hatchery_on_naturally_spawning_salmonids.pdf. Accessed July 25, 2012.
California Natural Resources Agency. 2010. Highlights of the BDCP. December 2010. Available at http://resources.ca.gov/docs/Highlights_of_the_BDCP_FINAL_12-14-10_2361.pdf. Accessed June 4, 2012.
Carr, G. M., H. C. Duthie, and W. D. Taylor. 1997 Models of aquatic plant productivity: A review of the factors that influence growth. Aquatic Botany 59:195-215.
CDFG (California Department of Fish and Game). 1996. Steelhead Restoration and Management Plan for California. The Resources Agency. Sacramento, CA.
CDFG. 1998. A status review of the spring run Chinook salmon in the Sacramento River drainage. Report to the Fish and Game Commission. Candidate species status report 98-1. Sacramento, CA.
CDFG. 2004. Sacramento River winter-run Chinook salmon biennial report 2002-2003. Prepared for the Fish and Game Commission by CDFG.
CDFG. 2009. Bay-Delta Sport Fishing Enhancement Stamp 2008/2009 Fiscal Year Fund Report. CDFG. Sacramento, CA.
CDFG and NMFS (National Marine Fisheries Service). 2001. Final Report on Anadromous Salmonid Fish Hatcheries in California. Joint Hatchery Review Committee. Available at http://nrm.dfg.ca.gov/documents/ContextDocs.aspx?cat=Fisheries. Accessed July 25, 2012.
Christie, M. R., M. L. Marine, R. A. French, and M.S. Blouin. 2012. Genetic adaptation to captivity can occur in a single generation. Proceedings of the National Academy of Sciences of the United States of America 109:238-242. Available at http://www.pnas.org/content/109/1/238.short. Accessed June 4, 2012.
Chung, F., and M. Ejeta. 2011 Estimating California Central Valley Unimpaired Flows. Presentation to California State Water Resources Control Board, January 11.
Clark, G. H. 1929. Sacramento-San Joaquin salmon (Oncorhynchus tshawytscha) fishery of California. California Fish and Game Bulletin 17:1-73.
Cloern, J. E. 1982. Does the benthos control phytoplankton biomass in south San Francisco Bay (USA)? Marine Ecology-Progress Series 9:191-202.
Cloern, J. E. 2001. Our evolving conceptual model of the coastal eutrophication problem. Marine Ecology Progress Series 210:223-253.
Cloern, J. E. 2007. Habitat connectivity and ecosystem productivity: Implications from a simple model. American Naturalist 169(1):E21-E33.
Cloern, J. E., and R. Dufford. 2005. Phytoplankton community ecology: Principles applied in San Francisco Bay. Marine Ecology Progress Series 285:1-28.
Cloern, J. E., K. A. Hieb, T. Jacobson, B. Sanso, E. Di Lorenzo, M. Stacey, J. L. Largier, W. Meiring, W. T. Peterson, T. M. Powell, M. Winder, and A. D. Jassby. 2010. Biological communities in San Francisco Bay track large-scale climate forcing over the North Pacific. Geophysical Research Letters 37.
Cloern, J. E., N. Knowles, L. R. Brown, D. Cayan, M. D. Dettinger, T. L. Morgan, D. H. Schoellhamer, M. T. Stacey, M. van der Wegen, R. W. Wagner, and A. D. Jassby. 2011. Projected evolution of California’s San Francisco Bay-Delta-River system in a century of climate change. PLoS ONE 6(9).
Cloern, J. E., A. D. Jassby, J. Carstensen, W. A. Bennett, W. Kimmerer, R. Mac Nally, D. H. Schoellhamer, and M. Winder. 2012. Perils of correlating CUSUM-transformed variables to infer ecological relationships (Breton et al. 2006, Glibert 2010). Limnology and Oceanography 57(2):665-668.
Cohen, A. N., and J. T. Carlton. 1995. Non-indigenous aquatic species in a United States estuary: A case study of the biological invasions of the San Francisco Bay and Delta. Report prepared for the U.S. Fish and Wildlife Service. Washington, DC.
Cohen, A. N., J. T. Carlton, and M. C. Fountain. 1995. Introduction, dispersal and potential impacts of the green crab Carcinus maenas in San Francisco Bay, California. Marine Biology 122:225-237.
Cole, B. E., and J. E. Cloern. 1984. Significance of biomass and light availability to phytoplankton productivity in San Francisco Bay. Marine Ecology-Progress Series 15:15-24.
Cole, B. E., J. K. Thompson, and J. E. Cloern. 1992. Measurement of filtration rates by infaunal bivalves in a recirculating flume. Journal of Marine Research 113:219-225.
Collie, J. S., S. J. Hall, M. J. Kaiser, and I. R. Poiner. 2000. A quantitative analysis of fishing impacts on shelf-sea benthos. Journal of Animal Ecology 69:785-798.
Conklin, J. 2005. Dialogue Mapping: Building Shared Understanding of Wicked Problems. New York: Wiley.
Conley, D. J., H. W. Paerl, R. W. Howarth, D. F. Boesch, S. P. Seitzinger, K. E. Havens, C. Lancelot, and G. E. Likens. 2009. Controlling eutrophication: Nitrogen and phosphorus. Science 323:1014-1015.
Conover, D. O., S. A. Arnott, M. R. Walsh, and S. B. Munch. 2005. Darwinian fishery science: Lessons from the Atlantic silverside (Menidia menidia). Canadian Journal of Fisheries and Aquatic Sciences 62:730-737.
Cowen, R. K., C. B. Paris, and A. Srinivasan. 2006. Scaling of connectivity in marine populations. Science 311(5760):522-527, doi:10.1126/science.1122039.
Cramer, S. P., and D. B. Demko. 1997. The Status of Late-Fall and Spring Chinook Salmon in the Sacramento River Basin Regarding the Endangered Species Act. Special Report submitted to the National Marine Fisheries Service on behalf of the Association of California Water Agencies and California Urban Water Agencies. Sacramento, CA.
Culberson, S., L. Bottorff, M. Roberson, and E. Soderstrom. 2008. Geophysical Setting and Consequences of Management in the Bay-Delta. Pp. 37-54 in The State of the Bay-Delta Science, M. C. Healey, M .D. Dettinger, and R. B. Norgaard, eds. The CALFED Science Program: Sacramento, CA.
D’Elia, C. F. 1987. Nutrient enrichment of the Chesapeake Bay: Too much of a good thing. Environment 29:2-10.
Davis, J., J. L. Grenier, D. Yee, L. J. McKee, B. K. Greenfield, R. Looker, C. Austin, M. Marvin-DiPasquale, R. Brodberg, and J. D. Blum. 2011. Reducing Methylmercury Accumulation in the Food Webs of San Francisco Bay and Its Local Watersheds. Environmental Research.
Denman, K., Y. Njiri, and H. Portner. 2010. 2010 Sendai ocean acidification workshop. Newsletter of the North Pacific Marine Science Organization 18(2):18-19.
Dettinger, M. D., and D. R. Cayan. 1995: Large-scale atmospheric forcing of recent trends toward early snowmelt runoff in California. Journal of Climate 8:606-623.
DiLorenzo, E., and S. Minobe. 2010. 2009 Mechanism of North Pacific Low Frequency Variability Workshop. Newsletter of the North Pacific Marine Science Organization 18(2):12-14.
DiLorenzo, E., J. E. Keister, V. Combes, and H. Batchelder. 2010. The Pacific Ocean Boundary Ecosystem and Climate Study (POBEX). Newsletter of the North Pacific Marine Science Organization 18(2):40-43.
Dugdale, R. C., F. P. Wilkerson, V. E. Hogue, and A. Marchi. 2007. The role of ammonium and nitrate in spring bloom development in San Francisco Bay. Estuaries 73(1-2):17-29.
Elmgren, R., and U. Larsson. 2001. Nitrogen and the Baltic Sea: Managing nitrogen in relation to phosphorus. The Scientific World 1(S2):371-377.
Feijoo, C. S., F. R. Momo, C. A. Bonetto, and N. M. Tur. 1996. Factors influencing biomass and nutrient content of the submersed macrophyte Egeria densa planch. in a pampasic stream. Hydrobiologia 341:21-26.
Feijoo, C., M. E. Garcia, F. Momo, and J. Toja. 2002. Nutrient absorption by the submerged macrophyte Egeria densa planch.: Effect of ammonium and phosphorus availability in the water column on growth and nutrient uptake. Limnetica 21:93-104.
Feyrer, F. 2004. Ecological segregation of native and alien larval fish assemblages in the southern Sacramento–San Joaquin Delta. American Fisheries Society Symposium 39:67-79.
Feyrer, F., B. Herbold, S. A. Matern, and P. B. Moyle. 2003. Dietary shifts in a stressed fish assemblage: Consequences of a bivalve invasion in the San Francisco Estuary. Environmental Biology of Fishes 67:277-288.
Feyrer, F., T. Sommer, and W. Harrell. 2006. Managing floodplain inundation for native fish: Production dynamics of age-0 splittail (Pogonichthys macrolepidotus) in California’s Yolo Bypass. Hydrobiologia 573:213-226.
Feyrer, F., M. L. Nobriga, and T. R. Sommer. 2007. Multi-decadal trends for three declining fish species: Habitat patterns and mechanisms in the San Francisco Estuary, California, USA. Canadian Journal of Fisheries and Aquatic Sciences 64:723-734. Available at http://www.water.ca.gov/aes/docs/FeyrerNobrigaSommer2007.pdf. Accessed July 18, 2012.
Fischer, H. B., E. J. List, J. Imberger, R. C. Y. Koh, and N. H. Brooks. 1979. Mixing in Inland and Coastal Waters. New York: Academic Press.
Fisher, T. R., E. R. Peele, J. A. Ammerman, and L. W. Harding. 1992. Nutrient limitation of phytoplankton in Chesapeake Bay. Marine Ecology Progress Series 82:51-63.
Fleenor, W. E., W. A. Bennett, P. B. Moyle, and J. R. Lund. 2010. On Developing Prescriptions for Freshwater Flows to Sustain Desirable Fishes in the Sacramento-San Joaquin Delta. Technical Report, Center for Watershed Sciences, University of California, Davis, 46 pp.
Foott, J. S., D. Free, T. McDowell, K. Arkush, and R. P. Hedrick. 2006. Infectious hematopoietic necrosis virus transmission and disease among juvenile Chinook salmon exposed in culture compared to environmentally relevant conditions. San Francisco Estuary and Watershed Science 4(1). Available at http://escholarship.org/uc/item/4cq1g6gd. Accessed July 17, 2012.
Foott, J. S., K. True, and R. Stone. 2008. Histological Evaluation and Viral Survey of Juvenile Longfin Smelt (Spirinchus thaleichthys) and Threadfin Shad (Dorosoma petenense) collected from the Sacramento – San Joaquin River Delta, April – November 2007. USFWS Ca-NV Fish Health Center, Anderson, CA. Available at http://www.fws.gov/canvfhc, Activities / reports.
Fox, J. P. 1987. Freshwater Inflow to San Francisco Bay under Natural Conditions. State Water Contracts Exhibit 262. California State Water Resources Hearings on the Bay-Delta. Sacramento, CA.
Geyer, W. R., 1993. The importance of suppression of turbulence by stratification on the estuarine turbidity maximum. Estuaries 16:113-125.
Glibert, P. M. 2010. Long-term changes in nutrient loading and stoichiometry and their relationships with changes in the food web and dominant pelagic fish species in the San Francisco Estuary, California. Reviews in Fisheries Science 18:211-232.
Glibert, P. M., D. Fullerton, J. M. Burkholder, J. C. Cornwell, and T. M. Kana. 2011. Ecological stoichiometry, biogeochemical cycling, invasive species, and aquatic food webs: San Francisco Estuary and comparative systems. Reviews in Fisheries Science 19(4):358-417.
Good, T. P., R. S. Waples, and P. Adams. 2005. Updated Status of Federally Listed ESUs of West Coast Salmon and Steelhead. U.S. Department of Commerce, NOAA Technical Memorandum NMFS-NWFSC-66.
Greene, V. E., L. J. Sullivan, J. K. Thompson, and W. J. Kimmerer. 2011. Grazing Impact of the invasive clam Corbula amurensis on the microplankton assemblage of the northern San Francisco Estuary. Marine Ecology Progress Series 431:183-193.
Greenfield, B. K., J. A. Davis, R. Fairey, C. Roberts, D. Crane, and G. Ichikawa. 2005. Seasonal, interannual, and long-term variation in sport fish contamination, San FranciscoBay. Science of the Total Environment 336:25-43.
Grimaldo, L. F., R. E. Miller, C. M. Peregrin, and Z. P. Hymanson. 2004. Spatial and temporal distribution of native and alien ichthyoplankton in three habitat types of the Sacramento–San Joaquin Delta. American Fisheries Society Symposium 39:81-96.
Grimaldo, L. F., A. R. Stewart, and W. Kimmerer. 2009a. Dietary segregation of pelagic and littoral fish assemblages in a highly modified tidal freshwater estuary. Marine and Coastal Fisheries: Dynamics, Management, and Ecosystem Science 1:200-217.
Grimaldo, L. F., T. Sommer, N. Van Ark, G. Jones, E. Holland, P. Moyle, B. Herbold, and P. Smith. 2009b. Factors affecting fish entrainment into massive water diversions in a freshwater tidal estuary: Can fish losses be managed? North American Journal of Fisheries Management 29:1253-1270.
Gross, E. S., J. R. Koseff, and S. G. Monismith. 1999. Three-dimensional salinity simulations in South San Francisco Bay. Journal of the Hydraulics Division ASCE 125(11):1199-1209.
Hallock, R. J,. W. F. Van Woert, and L. Shapovalov. 1961. Fish Bulletin No. 114. An Evaluation of Stocking Hatchery-Reared Steelhead Rainbow Trout (Salmo gairdnerii gairdnerii) in the Sacramento River System. eScholarship, University of California.
Healey, M. 2008. Science and the Bay-Delta. Pp. 19-35 in The State of Bay-Delta Science: 2008, edited by M. Healey. Sacramento, CA: CALFED Science Program. Available at http://www.science.calwater.ca.gov/publications/sbds.html. Accessed July 17, 2012.
Healey, M., M. Dettinger, and R. Norgaard. 2008a. New perspectives on science and policy in the Bay-Delta. Pp. 1-18 in The State of Bay-Delta Science: 2008, edited by M. Healey. Sacramento, CA: CALFED Science Program. Available at http://www.science.calwater.ca.gov/publications/sbds.html. Accessed July 17, 2012.
Healey, M. C., M. D. Dettinger, and R. B. Norgaard, eds. 2008b. The State of Bay-Delta Science, 2008. Sacramento, CA: CALFED Science Program.
Heinz Center. 2002. Dam Removal: Science and Decision Making. Washington, DC: The John Heinz III Center for Science, Economics, and the Environmnent.
Heppell, S. S. 2007. Elasticity analysis of green sturgeon life history. Environmental Biology of Fishes 79:357-368.
Herren, J. R., and S. S. Kawasaki. 2001. Inventory of water diversions in four geographic areas in California’s Central Valley. Pp. 343-355 in Contributions to the Biology of Central Valley Salmonids, edited by R. L. Brown. Volume 2. California Fish and Game, Fish Bulletin 179.
Heublein, J. C., J. T. Kelly, C. E. Croker, A. P. Klimley, and S. T. Lindley. 2009. Migration of green sturgeon, Acipenser medirostris in the Sacramento River. Environmental Biology of Fishes 84:245-258.
Hillman, T. W., and J. W. Mullan. 1989. Effect of hatchery releases on the abundance and behavior of wild juvenile salmonids. Pp. 265-285 in Summer and Winter Ecology of Juvenile Chinook Salmon and Steelhead Trout in the Wenatchee River, Washington, edited by Don Chapman Consultants. Final Report to Chelan County Public Utility District.
Hoekstra, J. M., K. K. Bartz, M. H. Ruckelshaus, J. M. Moslemi, and T. K. Harms. 2007. Quantitative threat analysis for management of an imperiled species: Chinook salmon (Oncorhynchus tshawytscha). Ecological Applications 17:2061-2073.
Hornberger, M.I., S. N. Luoma, A. van Geen, C. Fuller, and R. Anima. 1999. Historical trends of metals in the sediments of San Francisco Bay, California. Marine Chemistry 64:39-55.
Hornberger, M. I., S. N. Luoma, D. J. Cain, F. Parchaso, C. L. Brown, R. M. Bouse, C. Wellise, and J. K. Thompson. 2000. Linkage of bioaccumulation and biological effects to changes in pollutant loads in South San Francisco Bay. Environmental Science Technology 34:2401-2409.
HSRG (Hatchery Scientific Review Group). 2009. System-Wide Report on Columbia River Hatchery Reform. Available at http://hatcheryreform.us/hrp/reports/system/welcome_show.action Accessed July 16, 2012.
Hunt, J. W., B. S. Anderson, B. M. Phillips, R. S. Tjeerdema, K. M. Taberski, C. J. Wilson, H. M. Puckett, M. Stephenson, R. Fairey, and J. Oakden. 2001. A large-scale categorization of sites in San Francisco Bay, USA, based on the sediment quality triad, toxicity identification evaluations, and gradient studies. Environmental Toxicology and Chemistry 20:1252-1265.
IEP (Interagency Ecological Program for the San Francisco Estuary). 2006. Interagency Ecological Program Synthesis of 2005 Work to Evaluate the Pelagic Organism Decline (POD) in the Upper San Francisco Estuary. Available at http://www.water.ca.gov/iep/docs/pod/synthesis_report_111405.pdf. Accessed July 25, 2012.
Ihde, T. F., M. J. Wilberg, D. L. Loewensteiner, D. H. Secor, and T. J. Miller. 2011. The increasing importance of marine recreational fishing in the US: Challenges for management. Fish and Fisheries 108:268-276.
Ingram, B. L., J. C. Ingle, and M. E. Conrad. 1996. A 2000 yr record of Sacramento-San Joaquin River inflow to San Francisco Bay estuary, California. Geology 24:331-334.
Irvine, J. 2010. 2010 Sendai Salmon Workshop on Climate Change. Newsletter of the North Pacific Marine Science Organization 18(2):22-23.
Israel, J. A., K. J. Bando, E. C. Anderson, and B. May. 2009. Polyploid microsatellite data reveal stock complexity among estuarine North American green sturgeon (Acipenser medirostris). Canadian Journal of Fisheries and Aquatic Sciences 66:1491-1504.
Israel, J. A., and A. P. Klimley. 2008. Life history conceptual model for North American green sturgeon (Acipenser medirostris). In Delta Regional Ecosystem Restoration Implementation Plan. University of California, Davis. Available at http://nrm.dfg.ca.gov/documents/ContextDocs.aspx?sub=DRERIP_Documents_Models. Accessed July 25, 2012.
Israel, J. A., and B. May. 2010. Indirect genetic estimates of breeding population size in the polyploid green sturgeon (Acipenser medirostris). Molecular Ecology 19:1058-1070.
Iwama, G., and T. Nakanishi. 1996. The fish immune system, organism, pathogen, and environment. Fish Physiology 15.
Jackson, J. B. C., M. X. Kirby, W. H. Berger, K. A. Bjorndal, L. W. Botsford, B. J. Bourque, R. H. Bradbury, R. Cooke, J. Erlandson, J. A. Estes, T. P. Hughes, S. Kidwell, C. B. Lange, H. S. Lenihan, J. M. Pandolfi, C. H. Peterson, R. S. Steneck, M. J. Tegner, and R. R. Warner. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science 293:629-637.
Jaffe, B. E., R. E. Smith, and L. Z. Torresan. 1998. Sedimentation and bathymetric change in San Pablo Bay: 1856-1983. U.S. Geological Survey Open File Report 98-759. Sacramento, CA.
Janssen, E. M.-L., J. K. Thompson, S. N. Luoma, and R. G. Luthy. 2011. PCB-induced changes of a benthic community and expected ecosystem recovery following in situ sorbent amendment. Environmental Toxicology and Chemistry 30(8):1819-1826.
Jassby, A. D. 2008. Phytoplankton in the Upper San Francisco Estuary: Recent biomass trends, their causes and their trophic significance. Sacramento-San Joaquin Delta. San Francisco Estuary and Watershed Science 6(2).
Jassby, A. D., J. E. Cloern, and B. E. Cole. 2002. Annual primary production: Patterns and mechanisms of change in a nutrient-rich tidal ecosystem. Limnology and Oceanography 47:698-712.
Jassby, A. D., W. M. Kimmerer, S. G. Monismith, C. Armor, J. E. Cloern, T. M. Powell, J. Schubel, and T. Vendlinski. 1995. Isohaline position as a habitat indicator for estuarine resources: San Francisco Bay-Delta, California, U.S.A. Ecological Applications 5:273-289.
Jassby, A. D., and T. M. Powell. 1994. Hydrodynamic influences on interannual chlorophyll variability in an estuary: Upper San Francisco Bay-Delta (California, U.S.A.). Estuarine, Coastal, and Shelf Science 39:595-618.
Johnson, M., I. Werner, S. Teh, and F. Loge. 2010. Evaluation of Chemical, Toxicological and Histopathological Data to Determine Their Role in the Pelagic Organism Decline. Report for the Interagency Ecological Program.
Kimmerer, W. J. 2002. Effects of freshwater flow on abundance of estuarine organisms: physical effects or trophic linkages? Marine Ecology Progress Series 243:39-55.
Kimmerer, W. J. 2004. Open water processes of the San Francisco Estuary: From physical forcing to biological responses. San Francisco Estuary and Watershed Science 2(1).
Kimmerer, W. J. 2006. Response of anchovies dampens effects of the invasive bivalve Corbula amurensis on the San Francisco Estuary foodweb. Marine Ecology Progress Series 324:207-218.
Kimmerer, W. J. 2008a. Investigating particle transport and fate in the Sacramento-San Joaquin Delta using a particle tracking model. San Francisco Estuary and Watershed Science 6(1).
Kimmerer, W. J. 2008b. Losses of Sacramento River Chinook salmon and delta smelt to entrainment in water diversions in the Sacramento-San Joaquin Delta. San Francisco Estuary and Watershed Science 6(2):Article 2. Available at http://escholarship.org/uc/item/7v92h6fs. Accessed March 15, 2010.
Kimmerer, W. J. 2011. Modeling Delta Smelt Losses at the South Delta Export Facilities. San Francisco Estuary and Watershed Science 9(1). Available at http://escholarship.org/uc/item/0rd2n5vb#page-1. Accessed July 17, 2012.
Kimmerer, W. J., J. R. Burau, and W. A. Bennett. 1998. Tidally-oriented vertical migration and position maintenance of zooplankton in a temperate estuary. Limnology and Oceanography 43:1697-1709.
Kimmerer, W. J., E. S. Gross, and M. L. MacWilliams. 2009. Is the response of the estuarine nekton to freshwater flow in the San Francisco Estuary explained by variation in habitat volume? Estuaries and Coasts 32:375-389, doi:10.1007/s12237-008-9124-x.
Kimmerer, W. J., and M. L. Nobriga. 2008. Investigating particle transport and fate in the Sacramento-San Joaquin Delta using a particle tracking model. San Francisco Estuary and Watershed Science 6(1): Article 4.
Kimmerer, W. J., M. L. Nobriga, and L. Matthew. 2008. Investigating particle transport and fate in the Sacramento-San Joaquin Delta using a particle transport model. San Francisco Estuary and Watershed Science 6(1).
Kitchell, J. F., S. J. D. Martell, C. J. Walters, O. P. Jensen, I. C. Kaplan, J. Watters, T. E. Essington, and C. H. Boggs. 2006. Billfishes in an ecosystem context. Bulletin of Marine Science 79:669-682.
Koseff, J. R., J. K. Holen, S. G. Monismith, and J. E. Cloern. 1993. The effects of vertical mixing and benthic grazing on phytoplankton populations in shallow turbid estuaries Journal of Marine Research 51(4):1-26.
Krone, R. B. 1979. Sedimentation in the San Francisco Bay system. Pp. 85-95 in San Francisco Bay: The Urbanized Estuary, edited by T. J. Conomos. San Francisco, CA: Pacific Division, American Association for the Advancement of Science.
Kuivila, K. M., and M. L. Hladik. 2008. Understanding the occurrence and transport of current-use pesticides in the San Francisco Estuary Watershed. San Francisco Estuary and Watershed Science 6(3):Article 2. Available at http://repositories.cdlib.org/jmie/. Accessed July 17, 2012.
Lampman, B.-H. 1946. The Coming of the Pond Fishes. Portland, OR: Binford and Morts.
Laprise, R., and J. J. Dodson. 1994. Environmental variability as a factor controlling spatial patterns in distribution and species diversity of zooplankton in the St-Lawrence estuary. Marine Ecology Progress Series 107:67-81.
Lehman, P. W. 2010. The influence of climate on phytoplankton community biomass in San Francisco Bay Estuary. Limnology and Oceanography 45:580-590.
Lehman, P. W., G. Boyer, C. Hall, S. Waller, and K. Gehrts. 2005. Distribution and toxicity of a new colonial Microcystis aeruginosa bloom in the San Francisco Bay Estuary, California. Hydrobiologia 541:87-99.
Lehman, P. W., G. Boyer, M. Satchwell, and S. Waller. 2008. The influence of environmental conditions on the seasonal variation of Microcystis cell density and microcystins concentration in San Francisco Estuary. Hydrobiologia 600(1):187-204.
Lehman, P. W., S. J. The, G. L. Boyer, M. L. Nobriga, E. Bass, and C. Hogle. 2010. Initial impacts of Microcystis aeruginosa blooms on the aquatic food web in the San Francisco Estuary. Hydrobiologia 637:229-248.
Lindley, S. T., R. Schick, B. P. May, J. J. Anderson, S. Greene, C. Hanson, A. Low, D. McEwan, R. B. MacFarlane, C. Swanson, and J. G. Williams. 2004. Population structure of threatened and endangered Chinook salmon ESU in California’s Central Valley basin. NMFS Southwest Science Center NOAA-TM-NMFS-SWFSC-360. Santa Cruz, CA.
Lindley, S. T., R. Schick, A. Agrawal, M. Goslin, T. Pearson, E. Mora, J. J. Anderson, B. May, S. Greene, C. Hanson, A. Low, D. McEwan, R. B. MacFarlane, C. Swanson, and J. G. Williams. 2006. Historical population structure of Central Valley steelhead and its alteration by dams. San Francisco Estuary and Watershed Science 4(1):Article 3, 1-19. Available at http://repositories.cdlib.org/jmie/sfews/vol4/iss1/art3. Accessed July 17, 2012.
Lindley, S. T., R. S. Schick, E. Mora, P. B. Adams, J. J. Anderson, S. Greene, C. Hanson, B. P. May, D. McEwan, R. B. MacFarlane, C. Swanson, and J. G. Williams. 2007. Framework for Assessing Viability of Threatened and Endangered Chinook Salmon and Steelhead in the Sacramento-San Joaquin Basin. San Francisco Estuary and Watershed Science 5(1). Available at http://escholarship.org/uc/item/3653x9xc#page-2. Accessed July 25, 2012.
Lindley, S. T., M. L. Moser, D. L. Erickson, M. Belchik, D. W. Welch, E. L. Rechisky, J. T. Kelly, J. Heublein, and A. P. Klimley. 2008. Marine migration of North American green sturgeon. Transactions of the American Fisheries Society 137:182-194.
Lindley, S. T., C. B. Grimes, M. S. Mohr, W. Peterson, J. Stein, J. R. Anderson, L. W. Botsford, D. L. Botton, C. A. Busack, T. K. Collier, J. Ferguson, J. C. Garza, A. M. Grover, D. G. Hankin, R. G. Kope, P. W. Lawson, A. Low, R. B. MacFarlane, K. Moore, M. Plamer-Zwahlen, F. B. Schwing, J. Smith, C. Tracy, R. Webb, B. K. Wells, and T. H. Williams. 2009. What caused the Sacramento River fall Chinook stock collapse? National Marine Fisheries Service, Southwest Fisheries Science Center. NOAA_TM_SWFSC-447. Available at http://swr.nmfs.noaa.gov/media/salmondeclinereport.pdf. Acessed July 17, 2012.
Lluch-Coat, S. E., E. N. Curchitser, and S. Ito. 2010. 2010 Sendai Coupled Climate-to-Fish-to-Fisheries Models Workshop. Newsletter of the North Pacific Marine Science Organization 18(2):20-21.
Lodge, D. M., S. Williams, H. J. MacIsaac, K. R. Hayes, B. Leung, S. Reichard, R. N. Mack, P. B. Moyle, M. Smith, D. A. Andow, J. T. Carlton, and A. McMichael. 2006. Biological invasions: Recommendations for U.S. policy and management. Ecological Applications 16:2035-2054.
Lucas, L. 2009. Implications of estuarine transport for water quality. Pp. 273-307 in Contemporary Issues in Estuarine Physics, edited by A. Velle-Levinson. Cambridge: Cambridge University Press.
Lucas, L. V., J. E. Cloern, J. R. Koseff, S. G. Monismith, and J. K. Thompson. 1998. Does the Sverdrup critical depth model explain bloom dynamics in estuaries? Journal of Marine Research 56(2):375-415.
Lucas, L. V., J. E. Cloern, J. K. Thompson, and N. E. Monsen. 2002. Functional variability of habitats within the Sacramento-San Joaquin Delta: Restoration implications. Ecological Applications. 12: 1528-1547.
Lucas, L., J. R. Koseff, S. G. Monismith, and J. K. Thompson. 2009. Shallow water processes govern phytoplankton bloom dynamics: A modeling study. J. Mar. Systems 75:70-86.
Luoma, S. N., and J. E. Cloern. 1982. The impact of waste-water discharge on biological communities in San Francisco Bay. Pp137-160 in San Francisco Bay: Use and Protection, edited by W. J. Kockleman. Pacific Division of the AAAS. Available at http://sfbay.wr.usgs.gov/publications/pdf/luoma_1982_discharge.pdf. Accessed July 25, 2012.
Luoma, S. N., and D. J. H. Phillips. 1988. Distribution, variability and impacts of trace elements in San Francisco Bay. Marine Pollution Bulletin 19:413-425.
Luoma, S. N., and P. S. Rainbow. 2008. Metal Contamination in Aquatic Environments: Science and Lateral Management. Cambridge: Cambridge University Press.
Mac Nally, R., J. R. Thomson, W. J. Kimmerer, F. Feyrer, K. B. Newman, A. Sih, W. A. Bennett, L. Brown, E. Fleishman, S. D. Culberson, and G. Castillo. 2010. Analysis of pelagic species decline in the upper San Francisco Estuary using multivariate autoregressive modeling (MAR). Ecological Applications 20:1417-1430.
MacCready, P., and W. R. Geyer. 2010. Advances in estuarine physics. Annual Review of Marine Science 2:35-58, doi:10.1146/annurev-marine-120308-081015.
Maunder, M. N., and R. B. Deriso. 2011. A state-space multi-stage lifecycle model to evaluate population impacts in the presence of density dependence: Illustrated with application to delta smelt. Canadian Journal of Fisheries and Aquatic Sciences 68:1285-1306.
May, C., J. Koseff, L. Lucas, J. Cloern, and D. Schoellhamer. 2003. Effects of spatial and temporal variability of turbidity on phytoplankton blooms. Marine Ecology: Progress Series 254:111-128.
McDonald, E. T., and R. T. Cheng. 1997. A numerical model of sediment transport applied to San Francisco Bay, California. Journal of Marine Environmental Engineering 4:1-41.
McElhany, P., M. H. Ruckelshaus, M. J. Ford, T. C. Wainwright, and E. P. Bjorkstedt. 2000. Viable Salmonid Populations and the Recovery of Evolutionarily Significant Units. NOAA Techical Memorandum NMFS-NWFSC-42. National Marine Fisheries Service, NOAA, U.S. Department of Commerce.
McEwan, D. R. 2001. Central Valley steelhead. Pp. 1-43 in Contributions to the Biology of Central Valley Anadromous Salmonids, edited by R. Brown. Fish Bulletin 179, Volume 1. Sacramento, CA: California Department of Fish and Game.
Meyer, J. S., P. J. Mulholland, H. W. Paerl, and A. K. Ward. 2009. A Framework for Research Addressing the Role of Ammonia/Ammonium in the Sacramento-San Joaquin Delta and the San Francisco Bay Estuary Ecosystem. Report for the CALFED Science Program. Sacramento, CA: CALFED Science Program.
Michel, C. F., 2010. Losses of Sacramento River Chinook Salmon and Delta Smelt to Entrainment in Water Diversions in the Sacramento-San Joaquin Delta. Master’s thesis, University of California, Santa Cruz. 130 pp.
Miller, W.J. 2011. Revisiting assumptions that underlie estimates of proportional entrainment of delta smelt by State and Federal water diversions from the Sacramento-San Joaquin Delta. San Francisco Estuary and Watershed Science 9(1):4.
Miller, W. J., B. F. J. Manly, D. D. Murphy, D. Fullerton, and R. R. Ramey. 2012. An investigation of factors affecting the decline of delta smelt (Hypomesus transpacificus) in the Sacramento-San Joaquin Estuary. Reviews in Fisheries Science 20:1-19.
Mobrand, L., J. Barr, L. Blankenship, D. E. Campton, T. T. P. Evelyn, T. A. Flagg, C. V. Mahnken, L. W. Seeb, P. R. Seidel, and W. W. Smoker. 2005. Hatchery reform in Washington State: Principles and emerging issues. Fisheries June.
Modin, J. 1998. Whirling disease in California: A review of its history, distribution, and impacts, 1965-1997. Journal of Aquatic Animal Health 10(2):132-142.
Monismith, S. G., W. Kimmerer, J. R. Burau, and M. T. Stacey. 2002. Structure and flow-induced variability of the subtidal salinity field in Northern San Francisco Bay. Journal of Physical Oceanography 32:3003-3018.
Monismith, S. G., J. L. Hench, D. A. Fong, N. J. Nidzieko, W. E. Fleenor, L. Doyle, and S. G. Schladow. 2009. Thermal variability in a tidal river. Estuaries and Coasts 32(1):100-110, doi:10.1007/s12237-008-9109-9.
Monsen, N. E. 2000. A study of sub-tidal transport in Suisun Bay and the Sacramento–San Joaquin Delta, California. Ph.D. thesis, Stanford University.
Monsen, N. E., J. E. Cloern, and J. R. Burau. 2007. Effects of flow diversions on water and habitat quality: Examples from California‘s highly manipulated Sacramento-San Joaquin Delta. San Francisco Estuary and Watershed Science 5(3):Article 2, 1-13. Available at http://repositories.cdlib.org/jmie/sfews/vol5/iss3/art2/. Accessed July 17, 2012.
Mora, E. A., S. T. Lindley, D. L. Erickson, and A. P. Klimley. 2009. Do impassable dams and flow regulation constrain the distribution of green sturgeon in the Sacramento River, California? Journal of Applied Ichthyology 25:39-47.
Morse, H., and N. Manji. 2009. A thousand tiny wounds. Outdoor California 6-13.
Moyle, P. B. 2002. Inland Fishes of California, 2nd Edition. Berkeley, CA: University of California Press.
Moyle, P. B. 2008. The future of fish in response to large-scale change in the San Francisco Estuary, California. Pages 357-374 In K. D. McLaughlin, editor. Mitigating Impacts of Natural Hazards on Fishery Ecosystems. American Fishery Society, Symposium 64, Bethesda, Maryland.
Moyle, P., and J. Israel. 2005. Untested assumptions: Effectiveness of screening diversions for conservation of fish populations. Fisheries 30:20-28.
Moyle, P. B., J. R. Lund, W. A Bennett, and W. E. Fleenor. 2010. Habitat variability and complexity in the Upper San Francisco Estuary. San Francisco Estuary and Watershed Science 8(3).
Mueller-Solger, A. B., A. D. Jassby, and D. C. Muller-Navarra. 2002. Nutritional quality of food resources for zooplankton (Daphnia) in a tidal freshwater system (Sacramento–San Joaquin River Delta). Limnology and Oceanography 47:1468-1476.
Myers, R., J. Hutchings, and N. Barrowman. 1996. Hypotheses for the decline of cod in the North Atlantic. Marine Ecology Progress Series 138:293-308.
Newman, K. B. 2003. Modeling paired release-recovery data in the presence of survival and capture heterogeneity with application to marked juvenile salmon. Statistical Modeling 3:157-177.
Newman, K. B. 2008. An evaluation of four Sacramento-San Joaquin River Delta juvenile salmon survival studies. Stockton, CA: U.S. Fish and Wildlife Service.
Newman, K. B., and J. Rice. 2002. Modeling the survival of Chinook salmon smolts outmigrating through the lower Sacramento River system. Journal of the American Statistical Association 97:983-993.
Nichols, F., J. Cloern, S. Luoma, and D. Peterson. 1986. The modification of an estuary. Science 231:567-573.
Nichols, F. H., J. K. Thompson, and L. E. Schemel. 1990. Remarkable invasion of San Francisco Bay (California, USA) by the Asian clam Potamocorbula amurensis. II. Displacement of a former community. Marine Ecology Progress Series 66:95-101.
Nixon, S. W. 1995. Coastal marine eturophication: A definition, social causes, and future concerns. Ophelia 41:199-219.
NMFS (National Marine Fisheries Service). 2009a. Biological Opinion on the Long-Term Central Valley Project and State Water Project Operations Criteria and Plan. Available at http://swr.nmfs.noaa.gov/ocap.htm. Accessed July 12, 2012.
NMFS. 2009b. Public Draft Recovery Plan for Sacramento River Winter-run Chinook Salmon and Central Valley Spring-run Chinook Salmon and the Distinct Population Segment of Central Valley Steelhead. Sacramento, CA: NMFS Southwest Regional Office.
Nobriga, M., and P. Cadrett. 2003. Differences among hatchery and wild steelhead: Evidence from Delta fish monitoring programs. Interagency Ecological Program for the San Francisco Estuary Newsletter 14(3):30?38.
Nobriga, M. L., and F. Feyrer. 2008. Diet composition in San Francisco Estuary striped bass: does trophic adaptability have its limits? Environmental Biology of Fishes 83(4):495-503, doi: 10.1007/s10641-008-9376-0.
Nobriga, M., and B. Herbold. 2009. The Little Fish in California’s Water Supply: A Literature Review and Life-History Conceptual Model for delta smelt (Hypomesus transpacificus) for the Delta Regional Ecosystem Restoration and Implementation Plan (DRERIP). Sacramento, CA: DRERIP.
North, E. W., and E. D. Houde. 2001. Retention of white perch and striped bass larvae: Biological-physical interactions in the Chesapeake Bay turbidity maximum. Estuaries 24(5):756-769.
NRC (National Research Council). 1995. Science and the Endangered Species Act. Washington, DC: National Academy Press.
NRC. 1996. Upstream: Salmon and Society in the Pacific Northwest. Washington, DC: National Academy Press.
NRC. 1999. Sustaining Marine Fisheries. Washington, DC: National Academy Press.
NRC. 2004. Atlantic Salmon in Maine. Washington, DC: The National Academies Press.
NRC. 2006. Review of Recreational Fishery Survey Methods. Washington, DC: The National Academies Press.
NRC. 2010. A Scientific Assessment of the Alternatives for Reducing Water Management Effects on Threatened and Endangered Fishes in California’s Bay-Delta. Washington, DC: The National Academies Press.
Orsi, J. J. 1986. Interaction between diel vertical migration of a mysidacean shrimp and two-layered estuarine flow. Hydrobiologia 137:79-87.
Orsi, J. J., T. E. Bowman, D. C. Marelli, and A. Hutchinson. 1983. Recent introduction of the planktonic calanoid copepod Sinocalanys doerrii (Centropagidae) from mainland China to the Sacramento-San Joaquin Estuary of California. Journal of Plankton Research 5: 357-375.
Ostrach, D. J., J. M. Low-Marchelli, K. J. Eder, S. J. Whiteman, and J. G. Zinkl. 2008. Maternal transfer of xenobiotics and effects on larval striped bass in the San Francisco Estuary. Proceedings of the National Academy of Sciences of the United States of America 105(49):19354-19359.
Paerl, H. W. 1997. Coastal eutrophication and harmful algal blooms: Importance of atmospheric deposition and groundwater as “new” nitrogen and other nutrient sources. Limnology and Oceanography 42:1154-1165.
Paerl, H. W. 2008. Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater-marine continuum. Advances in Experimental Medicine and Biology 619:216-241.
Paerl, H. W. 2009. Controlling eutrophication along the freshwater–Marine Continuum: Dual nutrient (N and P) reductions are essential. Estuaries and Coasts 32:593-601.
Paerl, H. W., and J. Huisman. 2009. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environmental Microbiology Reports 1(1):27-37.
Paerl, H. W., R. S. Fulton, P. H. Moisander, and J. Dyble. 2001. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. The Scientific World 1:76-113.
Paerl, H. W., N. S. Hall, and E. S.Calandrino. 2011. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Science of the Total Environment 409:1739-1745.
Perry, R. W., J. R. Skalski, P. L. Brandes, P. T. Sandstrom, A. P. Klimley, A. Ammann, and B. MacFarlane. 2010. Estimating survival and migration route probabilities of juvenile chinook salmon in the Sacramento–San Joaquin River Delta. North American Journal of Fisheries Management 30(1):142-156.
Peterson, W., and K. Tadokoro. 2010. 2010 Sendai zooplankton workshop. PICES Press: Newsletter of the North Pacific Marine Science Organization 18(2):24-25.
PICES. 2010. Newsletter of the North Pacific Marine Science Organization 18(2).
Pine, W. E., I, S. J. D. Martell, C. J. Walters, and J. F. Kitchell. 2009. Counterintuitive responses of fish populations to management actions: Some common causes and implications for predictions based on ecosystem modeling. Fisheries 34:165-180.
Policansky, D. 1993. Fishing as a cause of evolution in fishes. Pp. 2-18 in The Exploitation of Evolving Resources, edited by T. K. Stokes, J. M. McGlade, and R. Law. Springer-Verlag. Berlin, Germany.
Presser, T. S. 1994. The Kesterson effect. Environmental Management 18:437-454.
Presser, T. S., and S. N. Luoma. 2006. Forecasting selenium discharges to the San Francisco Bay-Delta Estuary: Ecological effects of a proposed San Luis Drain extension. U.S. Geological Survey Professional Paper 1646. Available at http://pubs.usgs.gov/pp/p1646/. Accessed July 17, 2012.
Reynolds, C. S. 1987. Cyanobacterial water blooms. Advances in Botanical Research 13: 67-143.
Ricker, W. E. 1954. Stock and recruitment. Journal of the Fisheries Research Board of Canada 11:559-623. Res. Board Can. 11:559-623.
Ridderinkhof, H., and J. T. F. Zimmerman. 1992. Chaotic stirring in a tidal system. Science 2585085:1107-1109, doi:10.1126/science. 258.5085.1107.
Riley, S., H. Fuss, and L. LeClair. 2004. Ecological effects of hatchery-reared juvenile chinook and coho salmon on wild juvenile salmonids in two Washington streams. North American Journal of Fisheries Management 24:506-517.
Rittel, H., and M. Webber. 1973. Dilemmas in a general theory of planning. Policy Science 4:155-169.
Rjinsdorp, A. D. 1993. Selection differentials of male and female North Sea plaice Pleuronectes platessa L. and changes in maturation and fecundit. In The Exploitation of Evolving Populations, edited by R. Law, T. K. A. Stokes, and J. McGlade. London: Spring-Verlag.
Rose, K. A. 2000. Why are quantitative relationships between environmental quality and fish populations so elusive? Ecological Applications 10:367-385.
Ruhl, C., and D. Schoellhamer. 2004. Spatial and temporal variability of suspended-sediment concentrations in a shallow estuarine environment. San Francisco Estuary and Watershed Science 2(2).
Ruiz, G. M., J. T. Carlton, E. D. Grosholz, and A. H. Hines. 1997. Global invasions of marine and estuarine habitats by non-indigenous species: Mechanisms, extent, and consequences. American Zoologist 37:621-632.
San Joaquin River Group Authority. 2010. 2009 Annual Technical Report. P. 128 in On Implementation and Monitoring of the San Joaquin River Agreement and the Vernalis Adaptive Management Plan (VAMP). California Water Resources Control Board.
Schindler, D. W. 1971. Carbon, nitrogen, phosphorus and the eutrophication of freshwater lakes. Journal of Phycology 7:321-329.
Schoellhamer, D. H. 2000. Variability of suspended-sediment concentration at tidal to annual time scales in San Francisco Bay, USA. Continental Shelf Research 22: 1857-1866.
Schoellhamer, D. H. 2011. Sudden clearing of estuarine waters on crossing the threshold from transport- to supply- regulation of sediment transport as an erodible sediment pool is depleted: San Francisco Bay, 1999. Estuaries and Coasts 34:885-899.
Schreck, C. B. 1996. Inmmunomodulatin: Endogenous factors. Pp. 311-337 in The Fish Immune System, Volume 15: Organism, Pathogen, and Environment (Fish Physiology), edited by G. Iwama and T. Nakanishi. San Diego, CA: Academic Press.
Schwarzbach, S. E., J. D. Albertson, and C. M. Thomas. 2006. Effects of predation, flooding and contamination on reproductive success of California clapper rails (Rallus longirostris obsoletus) in San Francisco Bay. The Auk 123:45-60.
Simons, R., S. Monismith, F. Saucier, L. Johnson, and G. Winkler. 2007. Zooplankton retention in the estuarine transition zone of the St. Lawrence estuary. Limnology and Oceanography 51(6):2621-2631.
Smetacek, V., U. Bathmann, E.-M. Nöthig, and R. Scharek. 1991. Coastal eutrophication: Causes and consequences. Pp. 251-279 in Ocean Margin Processes in Global Change, edited by R. C. F. Mantoura, J.-M. Martin, and R. Wollast. Chichester: John Wiley and Sons.
Smith, S. E., and S. Kato. 1979. The fisheries of San Francisco Bay: Past, present and future. In 58th Annual Meeting of the Pacific Division of the American Association for the Advancement of Science. Pacific Division of the American Association for the Advancement of Science.
Snieszko, S. F. 1973. Recent advances in scientific knowledge and developments pertaining to diseases of fishes. Advances in Veterinary Science and Comparative Medicine 17:291-314.
Sommer, T., C. Armor, R. Baxter, R. Breuer, L. Brown, and M. Chotkowski. 2007. The collapse of pelagic fishes in the upper San Francisco Estuary. Fisheries 32(6):270-277.
Squire, S., G. H. Scelfo, J. Revenaugh, and A. R. Flegal. 2002. Decadal trends of silver and lead contamination in San Francisco Bay surface waters. Environmental Science and Technology 36:2379-2386.
Stacey, M. T. J. R. Burau, and S. G. Monismith. 2001. Creation of residual flows in a partially stratified estuary. Journal of Geophysical Research (Oceans) 106(C4):17013-17038.
Stewart, A. R., S. N. Luoma, C. E. Schlekat, M. A. Dobin, and K. A. Hieb. 2004. Food web pathway determines how selenium affects aquatic Ecosystems. Environmental Science & Technology 38:4519-4526.
Suchanek, T. H., C. A. Eagles-Smith, D. G. Slotton, E. J. Harner, and D. P. Adam. 2008. Mercury in abiotic matrices of Clear Lake, California: Human health and ecotoxicological implications. Ecological Applications 18(8 Suppl):A128-A157.
SWRCB (State Water Resources Control Board). 2010. Chapter 2: Beneficial Use in Water Quality Control Plan for the San Francisco Bay Basin. Available at http://www.swrcb.ca.gov/rwqcb2/water_issues/programs/planningtmdls/basinplan/web/docs/bp_ch2+tables.pdf. Accessed July 16, 2012.
Systech Water Resources, Inc. 2011. Task 5 Technical Memorandum; Link to CALSIM to Run WARMF Simulations. Available at http://www.waterboards.ca.gov/centralvalley/water_issues/drinking_water_policy/calsim_linkage.pdf. Accessed July 16, 2012.
Tamburri, M. N., K. Wasson, and M. Matsuda. 2002. Ballast water deoxygenation can prevent aquatic introductions while reducing ship corrosion. Biological Conservation 103:331-341.
Tatner, M. F. 1996. Natural chances in the immune system of fish. Pp. 255-287 in The Fish Immune System, Volume 15: Organism, Pathogen, and Environment (Fish Physiology), edited by G. Iwama and T. Nakanishi. San Diego, CA: Academic Press.
Thompson, J. 1957. Settlement Geography of the Sacramento–San Joaquin Delta, California. PhD dissertation, Stanford University.
Thompson, J. K. 2005. One estuary, one invasion, two responses: Phytoplankton and benthic community dynamics determine the effect of an estuarine invasive suspension-feeder. Pp. 291-316 in The Comparative Roles of Suspension-Feeders in Ecosystems, edited by R. F. Dame and S. Olenin. Amsterdam: Springer.
Thompson, J. K., and F. H. Nichols. 1996. Control of a phytoplankton bloom in San Francisco Bay, California by the filter feeding bivalve Potamocorbula amurensis [abstract]. Proceedings of the Pacific Division of the American Association for the Advancement of Science 13:98.
Thompson, J. K., J. R. Koseff, S. G. Monismith, and L. V. Lucas. 2008. Shallow water processes govern system-wide phytoplankton bloom dynamics: A field study. Journal of Marine Systems 74(1-2):153-166.
Thomson, J. R, W. J. Kimmerer, L. R. Brown, K. B. Newman, R. Mac Nally, W. A. Bennett, F. Feyrer, and E. Fleishman. 2010. Bayesian change-point analysis of abundance trends for pelagic fishes in the upper San Francisco Estuary. Ecological Applications 20:1431-1448.
USBR (U.S. Bureau of Reclamation). 2008. Biological Assessment of the Continued Long-Term Operation of Central Valley Project and State Water Project. Available at http://www.usbr.gov/mp/cvo/OCAP/sep08_docs/OCAP_BA_Aug08.pdf. Accessed July 16, 2012.
USBR. 2011. Reclamation Closing Red Bluff Diversion Dam Gates. Available at https://www.usbr.gov/newsroom/newsrelease/detail.cfm?RecordID=36494. Accessed July 17, 2012.
van Geen, A., and S. N. Luoma, eds. 1999. The impact of human activities on sediments of San Francisco Bay: An overview. Marine Chemistry. Special Issue 64:1-6.
Vogel, D. A., K. R. Marine, and J. G. Smith. 1988. Fish passage action program for Red Bluff Diversion Dam. Final Report on Fishery Investigations. Report No. FR1/FAO-88-19. Red Bluff, CA: U.S. Fish and Wildlife Service, Northern Central Valley Fishery Resource Office.
Vollenweider, R. A., R. Marchetti, and R. Viviani, eds. 1992. Marine Coastal Eutrophication. New York: Elsevier Science.
Wagner, R., M. T. Stacey, L. Brown, and M. Dettinger. 2011. Statistical models of temperature in the Sacramento-San Joaquin Delta under climate-change scenarios and ecological implications. Estuaries and Coasts 34:544-556.
Walters, R. A., R. T. Cheng, and T. J. Conomos. 1985. Time scales of circulation and mixing processes of San Francisco Bay waters. Hydrobiologia 129:13-36.
Waples, R. S. 1991. Genetic interactions between hatchery and wild salmonids—lessons from the Pacific Northwest. Canadian Journal of Fisheries and Aquatic Sciences 8:124-133.
Werner, I., F. G. Zalom, M. N. Oliver, L. A. Deanovic, T. S. Kimball, J. D. Henderson, B. W. Wilson, W. Krueger, and W. W. Wallender. 2004. Toxicity of storm-water runoff after dormant spray application in a french prune orchard, Glenn County, California, USA: Temporal patterns and the effect of ground covers. Environmental Toxicology and Chemistry 23(11):2719-2726.
Weston, D. P., and M. J. Lydy. 2010. Urban and agricultural sources of pyrethroid insecticides to the Sacramento-San Joaquin Delta of California. Environmental Science and Technology 44(5):1833-1840.
Weston, D. P., R. W. Holmes, J. You, and M. J. Lydy. 2005. Aquatic toxicity due to residential use of pyrethroid insecticides. Environmental Science and Technology 39(24):9778-9784.
Williams, J. G. 2006. Chapter 11: The Ocean. Pp. 216-231 in Central Valley Salmon: A Perspective on Chinook and Steelhead in the Central Valley of California. San Francisco Estuary and Watershed Science, 4(3). Available at http://escholarship.org/uc/item/21v9x1t7#page-216. Accessed July 16, 2012.
Williams, J. G. In press. Juvenile Chinook salmon (Oncorhynchus tshawytscha) in and around the San Francisco Estuary. San Francisco Estuary and Watershed Science.
Williams, S. L., and E. D. Grosholz. 2008. The invasive species challenge in estuarine and coastal environments: Marrying management and science. Estuaries & Coasts: Journal of the Coastal and Estuarine Research Federation 31:3-20.
Williams, P. B. and J. T. Hollibaugh. 1987. A Flow Standard to Maximize Phytoplankton Abundance by Positioning an Entrapment Zone in San Pablo Bay. State Water Resource Control Board.
Winder, M., and A. D. Jassby. 2011. Shifts in zooplankton community structure: Implications for food web processes in the upper San Francisco Estuary. Estuaries and Coasts 34:675-690.
Worm, B., E. B. Barbier, N. Beaumont, J. E. Duffy, C. Folke, B. S. Halpern, J. B. C. Jackson, H. K. Lotze, F. Micheli, S. R. Palumbi, E. Sala, K. A. Selkoe, J. J. Stachowicz, and R. Watson. 2006. Impacts of biodiversity loss on ocean ecosystem services. Science 314:787-790.
Wright, S. A., and D. H. Schoellhamer. 2004. Trends in the sediment yield of the Sacramento River, California, 1957–2001. San Francisco Estuary and Watershed Science 2(2).
Yemane, D., J. G. Field, and R. W. Leslie. 2005. Exploring the effects of fishing on fish assemblages using abundance biomass comparison (ABC) curves. ICES Journal of Marine Science 62:374-379.
Yoshiyama, R. M., E. R. Gerstung, F. W. Fisher, and P. B. Moyle. 1996. Historical and present distribution of Chinook salmon in the Central Valley drainage of California. Sierra Nevada Ecosystem Project: Final Report to Congress. Pp. 309-362 in Assessments, Commissioned Reports, and Background Information. Volume 3. Davis, CA: University of California, Center for Water and Wildland Resources, Davis CA.
































































































