
8
Social and Economic Underpinnings of Human Biodemography
Paul L. Hooper, Michael Gurven, and Hillard Kaplan
INTRODUCTION
The human species is an outlier in terms of demographic characteristics including lifespan, the duration of development, and menopause (Bogin and Smith, 1996; Gurven and Kaplan, 2007; Alberts et al., 2013). Human social relationships—including support from extended family, cooperative pair-bonds between men and women, biparental care, and extensive cooperation between non-relatives—are likewise features that set humans apart from many other primates and mammals (Lancaster and Lancaster, 1983; Hill et al., 2009; Hrdy, 2009; Jaeggi and Gurven, 2013). It is increasingly clear that patterns of human demography and sociality are interdependent and mutually reinforcing, making it likely that they co-evolved during humans’ evolutionary history. In this paper, we present a framework for investigating the relevant causal links between ecology, social structure, and biodemography that are essential to explaining the unique evolutionary trajectory of the species. This framework aims to explain both humans’ unique demographic and social characteristics relative to other taxa, as well as variation in these characteristics within and across human societies.
Our proposal is that three distinctive forms of social relationships evolved and were likely to have been present in most populations prior to the advent of agriculture: (1) kin-based altruism, with net downward transfers of food energy and other forms of assistance across generations; (2) complementary specialization of labor by men and women, and cooperation by couples in reproduction, childcare, household labor, and economic
production; and (3) cooperation and reciprocal assistance among related and unrelated individuals to buffer shortfalls and/or to coordinate efforts in food acquisition (see Figure 8-1). We posit that these social relationships are linked responses to the particular ecological niche to which humans are adapted. We also posit that those three classes of relationships have significant impacts on health and well-being throughout the lifecourse. As a result, they are likely to have played a critical role affecting natural selection on rates of aging and reproduction, and the ability to escape death from economic shortfalls, injury, or illness.
Given that we cannot directly observe the social relationships, demographic characteristics, and behavioral patterns of prehistoric populations during human evolution, we are reliant on data derived from the paleo-archeological record, primatology, and the study of contemporary human populations. The quantitative study of extant human groups engaged in foraging or a mix of foraging and rudimentary horticulture (forager-horticulturalists) in recent decades, in particular, has provided a more detailed understanding of the relationships between social, economic, and demographic variables under pre-modern circumstances than has previously been available. There have been both advantages and disadvantages of this approach.
The disadvantages derive from the fact that contemporary populations are not frozen relics of the past. In fact, it is always the case that by the
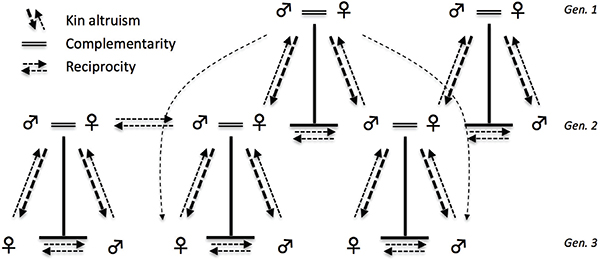
FIGURE 8-1 Evolved modal human social organization.
NOTE: The dominant pattern of relationships based on kin altruism, complementarity, and reciprocity are indicated by the dashed arrows and double solid lines: kinship motivates net downward flows of resources within families (vertical dashed arrows); complementarity promotes cooperative division of labor within pair bonds (double solid lines); and reciprocity supports bidirectional exchange within and between families (horizontal dashed lines).
time foraging populations are studied by scientists, their lifestyle and other cultural characteristics are undergoing change. Moreover, such populations are not randomly distributed in geographical space, but tend to live in remote areas that have had less potential for economic development. Therefore, they represent a nonrandom subset of the lifestyles that existed in places where developed states now dominate the landscape (Headland and Reid, 1991; Kelly, 1995). States and empires have also had the effect of repressing violence between small-scale groups, which has made it difficult to assess the impact of interpersonal and intergroup violence on life histories and behavior under pre-modern conditions (but see Patton, 2000; Wrangham et al. 2006; Hill et al., 2007; Sussman and Cloninger, 2011 for explicit treatments). Global environments have also changed in ways that are likely to have direct effects on the parameters of interest, particularly with respect to the distribution of plant and animal species important in forager diets, and pathogens affecting morbidity and mortality. Finally, all of the groups that have been extensively studied have experienced a unique set of historical forces in recent decades with differential impacts on current conditions and lifestyles (see, for example, Hill and Hurtado, 1996, on the Aché in Paraguay, compared with Headland, 1986, on the Agta in the Philippines).
The advantages derive from the fact it is possible to collect detailed, quantitative data on reproduction, morbidity, mortality, transfers of food and other forms of assistance, conflict, and social status in extant small-scale human communities. Age- and sex-specific patterns of fertility and mortality, work, food production and consumption, food sharing, childcare, and family relationships have been extensively studied in a sample of these populations in Africa, Australia, the Philippines, and South America. Our goal in this paper is to synthesize what appear to be the modal—and sometimes apparently universal—patterns in such groups within a coherent theoretical framework. The common patterns found in populations that have experienced such divergent histories in a variety of ecologies with no recent common cultural ties are likely to be robust features of humans while engaging in foraging-based lifestyles. Such common threads may also indicate their historical depth in human evolution.
In the following sections, we provide a characterization of modal patterns of social organization and demography among human foragers. For each of the key social relationships introduced above—kin-based altruism within families, cooperative pair-bonds, and reciprocal cooperation—we discuss insights derived from evolutionary economic models, and review available evidence from the study of modern human foragers and other small-scale human societies. We follow this with a characterization of variation within and away from modal patterns of social structure and demography among foragers and other preindustrial human groups. We
conclude by discussing the relevance of these patterns for understanding the evolutionary past and present behavior of the human species.
KIN-BASED ALTRUISM, INTERGENERATIONAL TRANSFERS, AND LIFE HISTORY
A crucial aspect of the economic system of human foragers is that production activities are skill-intensive and require a long investment process. Although both humans and chimpanzees (humans’ closest primate relative) are both omnivores, the composition of their diets is radically different. Data on diet from chimpanzees in the Gombe reserve in Tanzania indicate that about 95 percent of calories are derived from leaves and fruit. In contrast, for the sample of 10 human foraging societies for which quantitative data are available, fruits constitute an average of only 10 percent of the diet. The remaining 90 percent calories are derived from more difficult-to-acquire hunted and extracted (e.g., nuts, tubers, honey) foods, which are only about 5 percent of the chimpanzee diet (see Kaplan et al., 2000, for data sources).
The nature of the production and processing of foods has implications for how long it takes to become efficient at foraging and for the age at which juveniles become nutritionally independent. Efficiency in activities based on skill development tends to increase with age over a longer period compared with activities requiring less experience. This is pertinent for most traditional forms of hunting, as well as many skill-intensive methods of extraction and processing. The return rates of Aché and Tsimane hunters, for instance, continue to increase across the first three decades of life, and peak remarkably late, in the 30s and 40s, due to the importance of experience and skill development over physical strength alone (Walker et al., 2002; Gurven et al., 2006). The importance of skill-acquisition and learning also tends to reduce productivity early in life, as young invest time and energy in developing the brains, bodies, and skills necessary for later success. Children thus remain nutritionally dependent on parents until approximately 18-20 years of age among the foragers and forager-horticulturalists for which such data exist. Chimpanzee offspring, in contrast, begin to supplement the energy they receive from lactation with their own foraging efforts during their first or second year of life, and are largely energy self-sufficient by age 5 (Kaplan et al., 2000).
Figure 8-2 shows age-specific food production, consumption, and their net (production minus consumption) for Tsimane forager-horticulturalists averaged over males and females (Gurven et al., 2012). During infancy, childhood, and adolescence, individuals run large net deficits in production (more calories consumed than produced each day); during adulthood, however, individuals produce large net surpluses (more calories produced than
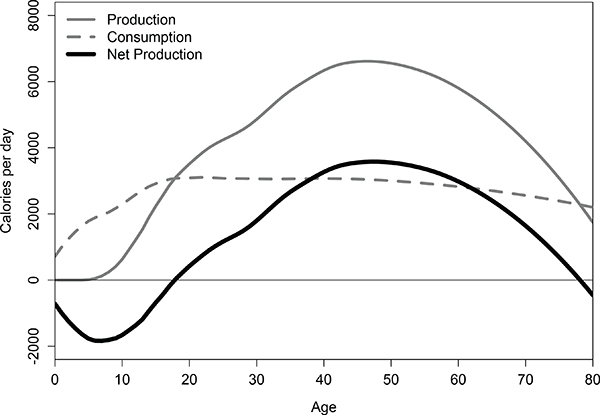
FIGURE 8-2 Mean production, consumption, and net production (production minus consumption) of food per day as a function of age among Tsimane forager-horticulturalists.
SOURCE: Gurven et al. (2012).
consumed), with a peak in net production in the beginning of middle age. Similar curves—with some variation in ages of peak production—have been reported for Aché, Hiwi, !Kung, and Hadza foragers, as well as for Piro and Machiguenga forager-horticulturalists (Gurven and Kaplan, 2006; Hill and Hurtado, 2009; Howell, 2010; Marlowe, 2010).1 As a result of the slow accumulation of human capital across life (particularly skills, experience, and ability), there are larger differences in productivity between young and old among humans than in other primates or mammals (an exception may be killer whales: Rendell and Whitehead, 2001; Foote, 2008).
Observed life cycles of production among hunter-gatherers are fundamentally interlinked with human biodemographic characteristics, including: (1) longevity; (2) intergenerational transfers; and (3) post-reproductive survival. Analytical models of life history evolution show that exogenous shifts
_______________
1Because most of these data are cross-sectional, secular trends may confuse age trends. It may be, for instance, that younger cohorts are healthier but less skilled at traditional activities; the former would tend to exaggerate apparent aging, while the latter would exaggerate the importance of lifelong learning.
in economic production to older ages due to learning result in selection for increased investment in survival (Kaplan and Robson, 2002; Robson and Kaplan, 2003). The evidence for the longevity of foragers under pre-modern conditions compared with wild chimpanzees has been reviewed in Gurven and Kaplan (2007). Available data show that conditional on surviving to age 15, a chimpanzee has an expectation of an additional 13 years of life, whereas foragers show an expectation of an additional 40 or so years of life (with an expected age of death in the later 50s). The contrast is even more extreme at age 45, when most women have ceased to reproduce: whereas a chimpanzee can expect to live less than an additional 5 years, a female forager can be expected to live an additional 22 years, to age 67. On average, the modal age of death (i.e., the age with the highest density of deaths) for those surviving to adulthood occurs sometime around age 70 in extant foraging populations.
Second, the age-schedules of productivity, mortality, and reproduction also affect the flow of resources between generations because they determine the distribution of individuals with caloric deficits and surpluses. Inclusive fitness theory provides a specific means of understanding transfers between kin as a function of differential productive ability and need (Hooper et al., 2014). This approach predicts that transfers are determined by the interaction of two factors: (1) the relative gains to the recipient compared with the costs to the donor, which are, in turn, determined by the productivity and consumption needs of individuals and families; and (2) their degree of kinship in terms of the likelihood of sharing genes by recent common descent. Early in life, when individuals are relatively inefficient producers and cannot meet their energy requirements through their own effort, the marginal benefit of receiving calories from others is higher. After efficiency increases at older ages, on the other hand, the marginal cost of giving away calories is reduced due to high productivity and diminishing returns to personal consumption. These effects are expected to interact positively with the degree of relatedness.
This logic provides a basis for understanding variation in parental investment, and net transfers of energy, time, and effort between kin more generally. In humans, asymmetries in productivity combine with the motivation to assist kin to produce systematic and predictable flows of resources from older to younger generations within families. Figure 8-3, adapted from Hooper (2011), illustrates the flow of food across three generations within Tsimane extended families. It plots the expectations of caloric transfers among older individuals (grandparental generation), their adult offspring, and grandchildren. The left-hand panel plots the gross transfers in either direction, while the right-hand panel presents the net transfers (gross transfer from individual i to individual j minus the gross transfer from j to i). The right-hand panel shows that net flows tend to be downward. The largest

FIGURE 8-3 Mean gross food transfers (left panel) and net food transfers (right panel) within Tsimane extended families.
SOURCE: Hooper (2011).
flows are within nuclear families, from parents to children, but there are also significant flows downward from older individuals to families of adult children and younger siblings. Among the Tsimane, mean transfers are net downward even through the seventh decade of life (Hooper et al., 2014).
While representations of transfers at this level of detail are not available in published data for other groups (but for the Hadza, see Hawkes et al., 1989, 1997; Wood and Marlowe, 2013), evidence suggests that transfers tend to flow among kin (reviewed in Gurven, 2004) and that downward transfers within families appear to be universal in all societies for which transfers have been measured (reviewed in Lee, 2007). In all observed cases, foragers remain net producers after they have ceased to reproduce, implying downward net transfers (Kaplan, 1994; Kaplan et al., 2000; Gurven and Kaplan, 2006; Hill and Hurtado, 2009; Howell, 2010; Marlowe, 2010). While food transfers outside lactation are much more limited in other primate species, sharing is more common for high-quality difficult-to-acquire foods and occurs principally from mothers to offspring (see Silk, 1978; Price and Feistner, 1993; Nishida and Turner, 1996; Jaeggi et al., 2008; Jaeggi and Van Schaik, 2011 for a cross-species analysis). Human foragers have taken this tendency to an outlying extreme.
Third, because transfers from older adults appear to support their adult children’s fertility and grandchildren, the life history of production and consumption also sheds light on the evolutionary economics of menopause (Williams, 1957; Lancaster and King, 1985; Hawkes et al., 1998; Kaplan et al., 2010). If all else were held constant but women continued to reproduce until older ages (e.g., age 55), lineages would find themselves in net caloric deficit. Either age-specific production rates or age-specific fertility rates would have to change (Kaplan et al., 2010). “Premature” reproductive cessation can then be thought of as an emphasis on economic contributions to younger generations at the cost of continued production of offspring. Given the similarities between chimpanzees and humans in age of menopause, it has been argued that selection may have favored lengthening of lifespan rather than menopause itself (see, for example, Ellison and Ottinger, this volume). In our view, both menopause and lifespan extension require explanation. Even if human-typical rates of reproductive decline were established prior to selection for lifespan extension in the human lineage, it is still necessary to explain why there would not be selection for continued reproductive function once ancestral people were living longer.
A final striking feature of the caloric transfers illustrated in Figure 8-3 is the economic role that men play, as husbands, fathers, and grandfathers. In general, there are positive net energy transfers from men to their wives, greater transfers to children from fathers than from mothers, and greater transfers to grandchildren from grandfathers than from grandmothers. To shed light on these patterns of investment, and their relationship with mating effort and mate choice, the next section addresses sex roles and mating systems in foraging societies.
COMPLEMENTARITIES, SEX ROLES, AND LIFE HISTORY
The evolution of patterns of mating and parental investment among humans can be understood in light of ecological factors underlying variation in animal mating systems more generally. For females (or males in sex-role reversed species), theory in evolutionary biology emphasizes fitness impacts of choice for male genetic quality versus paternal investment. For males, it focuses on fitness impacts of paternal investment versus investments in mating effort—for example, in displays of genetic quality or contests with other males. We propose that the relative weights of these fitness effects depends on the nature of tradeoffs between acquiring energy (i.e., food) and providing care to offspring in the time budgets of parents. They also depend on the degree of complementarity between investments of care and energy, and between these investments and genetic quality in determining offspring success (for relevant background and evolutionary-economic models, see Becker, 1973; Maynard Smith, 1977; Grafen and Sibly, 1978;
Lazarus, 1990; Clutton Brock, 1991; Kokko and Johnstone, 2002; Kokko and Jennions, 2008; Bowles and Borgerhoff Mulder, 2012).
In this framework, the fitness of offspring (W) can be considered a function of the inputs of genes (G), energy (E), and care (C) provided by female (f) and male (m) parents: W = w(Gf, Gm, Ef, Em, Cf, Cm). Complementarities in these inputs are reflected in the direction of their cross-partial derivatives. If the cross-partials are positive (i.e., W´GE > 0, W´GC > 0, or W´EC > 0), this reflects positive complementarities between inputs. For example, a positive cross-partial between genetic quality and energy implies that an increase in energy will have a larger fitness impact on an offspring of higher genetic quality, and vice versa. In the case of mammals, who provision their offspring through lactation, or birds, who provision offspring with food, there are likely to be especially strong complementarities between energy and care. A perfectly protected offspring who receives no energy will still likely die, as will a well-fed offspring that is not protected from predators. The problem facing each sex is to optimize these inputs in the face of a time budget constraint, T = t(tE,tC,tO), reflecting the division of time between acquiring energy, providing care, and other activities (such as mating effort).
We hypothesize that patterns of mating and parental investment respond to ecological conditions affecting complementarities and trade-offs between providing care and energy. Figure 8-4 depicts two different kinds of ecological conditions affecting the gains from biparental investment. Figure 8-4a represents what we hypothesize to be the modal mammalian pattern, with a convex budget constraint for individual parents (the solid black line) reflecting a relatively “easy” tradeoff between the production of care and energy. Here, a single parent is capable of providing both goods at a reasonably high level simultaneously, and increased specialization in either care or energy results in diminishing marginal returns.
Most (particularly, female) mammals engage in some strategy that allows for protecting young while foraging. This is accomplished by caching young in hidden burrows (practiced by most rodents), having them accompany on foot (practiced by most herbivores), or by carrying them (practiced by all primates). This means that, compared with the mother that combines care and energy acquisition, the mother who feeds exclusively at the expense of any protection will not get much more food, and the mother who exclusively protects at the expense of energy acquisition will not provide much more protection. These conditions reduce the gains from adding a second parent’s investments relative to what can be acheived by a single parent alone (compare points A and C in Figure 8-4a). This would hold especially where the type of foods that are consumed come in small, easy-to-acquire packages (like blades of grass or leaves) that are not efficiently shared. Under these conditions, we predict that the gains to females from choosing males with higher genetic quality will be greater than from choos-
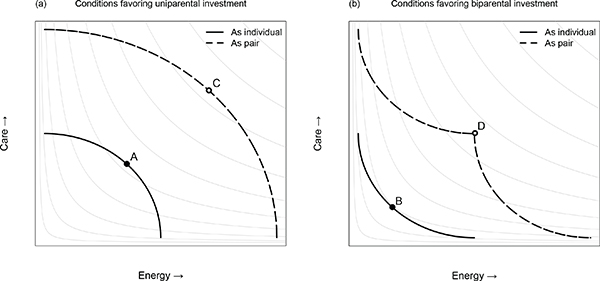
FIGURE 8-4 Hypothetical budget constraints reflecting tradeoffs between the provision of care and energy by parents to offspring.
NOTE: The solid black lines represent the feasible levels of care and energy which a single parent can provide, while the dashed black lines represent the levels of care and energy that two parents can provide together. The thin gray lines represent hypothetical iso-fitness lines for offspring, with increasing offspring fitness from the lower left to upper right of each panel. The solid points A and B represent the optimal allocation of effort for a single parent, while the open points C and D represent the optimal allocation of effort for two parents together. Panel (a) illustrates a convex budget constraint, reflecting diminishing returns to specialization in energy production or care. Under these conditions, because the fitness of offspring with two investing parents (point B) is less than twice that of offspring with a single parent (point A), one sex (usually the male) is likely to benefit from deserting to pursue additional mating opportunities, resulting in an equilibrium of uniparental investment at point A. Panel (b) illustrates a concave budget constraint, reflecting increasing returns to specialization or turn-taking by parents. Under these conditions, because the fitness of offspring with two parents dividing labor (point D) is more than twice that of offspring with a single parent (point C), the gains to biparental investment are likely to outweigh the benefits of desertion, resulting in an equilibrium of biparental investment with a specialized division of labor at point D. The curvature of the offspring fitness surface can also affect the equilibrium outcome, with a greater likelihood of biparental care when there are increasing returns to investment in offspring.
ing males who offer care or energy. This, in turn, affects the tradeoff for males between parental investment versus gaining access to females through competition, favoring the latter. These conditions are expected to foster provisioning of both care and energy by females, and higher mean quality of genes contributed by males than females (i.e., Gm > Gf).
Figure 8-4b depicts alternative ecological conditions, which we hypothesize to characterize birds emphasizing aerial locomotion, some species of fish, and humans. These conditions reflect a relatively harsher tradeoff between providing care versus energy and support biparental investment. This would occur, for example, where young are unable to accompany adults while foraging, but face high risks of predation if left unattended; where foraging activites would expose offspring to high levels of risk; or where foraging returns rates would greatly suffer if offspring accompanied adults. In these cases, the budget constraint defining the tradeoff between provisioning of care and energy for an individual parent will be tend to be concave (like the solid black line in Figure 8-4b), reflecting increasing returns to specialization in producing one good or the other at a time, but low returns to producing both at once. These conditions increase the relative gains to biparental over uniparental investment with a cooperative division of labor between parents (compare points B and D in Figure 8-4b). Mate choice under these conditions is expected to emphasize genetic quality relatively less, and parental investment offered and provided relatively more.
In most birds, eggs and nestlings are highly vulnerable to predation. Food supplies are often some distance from nests, not allowing for simultaneous care and energy acquisition. Different solutions to this problem appear to depend on the details of the local ecology, but often involve the help of a second parent (Bennett and Owens, 2002). In birds, a common solution is turn-taking, where one parent stays to protect nestlings while the other forages for food. Parental specialization also sometimes occurs, with one parent caring and the other provisioning. In many fish exhibiting biparental investment, females provide energy to offspring through investment in eggs, while males provide protective care by guarding the eggs from predation (Gross and Sargent, 1985). Males protect eggs and fry in safer places while females forage for energy where food supplies are rich. In other species, biparental mouthbrooding is common, which may allow parents to achieve returns to scale that are unattainable through the investment of one parent alone (Kidd et al., 2012).
The modal pattern among human foragers is characterized by life history specializations in care and gathering by females, and hunting by males. The returns to specialization turn on a number of factors. First, human mothers share with other primates a commitment to carrying infants and providing intensive maternal investment through lactation. Most hunting practices, on the other hand, are incompatible with pregnancy and childcare, given their danger and physicality. Given the skill-intensive nature of hunting and its incompatibility with pregnancy and childcare, women would be unlikely to attain the hunting return rates achieved by men due to the inability to invest sufficient time in practice (Kaplan et al., 2001).
Second, the returns to investment by both sexes are increased by the depth and duration of offspring need during development, as required by the brain- and experience-intensive nature of the human life history. (Similar effects of offspring need are observed in birds: Bennett and Owens, 2002.) Finally, the human pattern of multiple simultaneously dependent offspring further deepens the extent of economic need, drawing the investments of fathers and other close kin (Gurven and Walker, 2006).
The sexual division of labor is a ubiquitous feature of human societies—with the possible exception of some sectors of modern nations—that is well documented in the ethnographic literature (partially represented in the Human Relations Area files: Murdock and Provost, 1973). Among foragers, there is an extensive division of labor with respect to all aspects of household production, including not only direct care and energy production, but also food processing, cooking, collection of firewood, construction of housing, and production of tools (Marlowe, 2007). Compatibility of labor with simultaneous childcare is a strong predictor of women’s activities (Minge-Klevana, 1980; Hurtado et al., 1985). It is not that there is no overlap in the activity budgets of men and women, as men do engage in some direct care of infants, for example, among the Aché, Tsimane, !Kung, Aka, and most—perhaps all—other foragers and forager-horticulturalists (Hill and Hurtado, 1996; Winking et al., 2009; Howell, 2010; Hewlett, 1992; Hewlett and Macfarlan, 2010).
Among foragers, the weight of quantitative evidence suggests substantial contributions to familial and offspring well-being from fathers as well as mothers in the context of sex-specific specialization. Among the sample of 10 foraging societies for which quantitative data are available, males produce an average of 68 percent of all food calories and 88 percent of all protein (Kaplan et al., 2001). There is substantial variability across foraging societies in male and female energy inputs, but there are no societies reported for which men are not important providers of meat (with the possible exception of some aboriginal Australian groups for which quantitative data do not exist). The Tsimane show mean net caloric flows from husbands to wives, and net caloric contributions of fathers to offspring are roughly twice those of mothers (Figure 8-3; Hooper et al., 2014).
The behavior of female foragers reflects complementary adjustments to the investments of males, particularly in care and economic activities that are compatible with care. For example, among the Aché who did not maintain permanent camps with safe places for children, women with infants and toddlers under age 3 spent more than 90 percent of daytime hours in tactile contact with those children (Hurtado et al., 1985). Even in safer environments where cleared spaces are maintained, as among the Tsimane, children are actively cared for by their mothers about 30 percent of waking hours (Winking et al., 2009). Consisent with this emphasis on direct
care, female foragers show significant decreases in time spent foraging with the presence of young children. Among the Aché and Hiwi, women who are nursing show reduced time devoted to production and lower production rates than those who are not (Hurtado et al., 1985, 1992). Tsimane women’s rate of direct food production—at a mean base of 2,900 calories per day—is estimated to decrease by 900 calories for each child under age 3; it is estimated to increase, on the other hand, by 320 calories for each additional child over 3. On top of a mean base of 5,200 calories produced per day, Tsimane men also increase hunting effort with a greater number of dependents and produce an estimated 240 additional calories of animal fat and protein for each additional dependent offspring (Hooper et al., 2015; see also Marlowe, 2003, for the Hadza).
Complementarities between male and female parental roles affect patterns of marriage and reproduction among foragers in multiple ways. In a cross-cultural sample of 145 foraging groups, the modal percent of marriages that are polygynous is 0-4 percent, and more than 90 percent of marriages are monogamous in the majority of groups (Binford, 2001). Consistent with this pattern, for the foraging societies for which data exist, male and female reproductive schedules tend to be closely linked: Age-specific fertility and expected future fertility for women and men are similar in shape, with the male curves shifted 3-5 years to the right (i.e., to older ages) and having slightly longer tails (see Tuljapurkar et al., 2007, and original data from Howell, 1979; Melancon, 1982; Blurton Jones et al., 1992; Hill and Hurtado, 1996). This pattern differs from what has been characterized as the standard difference between the fertility schedules of male and female mammals. As male parental investment is rare among mammals, male-male competition tends to be high, leading to both later onset of reproduction and earlier termination of reproduction among males, compared with females (because it takes males longer to become competitive than for females to begin reproducing, and the effects of senescencing competitive abilities affects male reproduction more than female; Clutton-Brock, 1991; Clutton-Brock and Isvaran, 2007; see also Alberts et al., this volume).
The demographic linkage between women and men is also manifest in the tendency for male foragers to cease reproducing when their wives reach menopause. In the case of Tsimane, only 10 percent of men reproduce after their wife reaches menopause (Kaplan et al., 2010). Similarly, among the Aché about 17 percent of men reproduced after their wife reached menopause, and only 10 percent reproduced if they had two or more children with that wife (Hill and Hurtado, 1996). As depicted in Figure 8-3, as their wives reach menopause, Tsimane men tend to invest in their children and their grandchildren rather than investing in new reproductive careers. Older men in foraging societies may not be very attractive as mates for young
women. Given that children will be dependent for a long time, men whose productive abilities are on the wane do not offer good long-term prospects. Moreover, ceteris paribus, it is likely that women would prefer to have all their children with one man, since serial fathers would be differentially related to the children (Kaplan et al., 2010).
GAINS TO COOPERATION, RECIPROCITY, AND LIFE HISTORY
In addition to the intensive relationships within nuclear and extended families, cooperation between more distant kin and non-kin is another hallmark of human sociality. This can be defined in terms of both the breadth of relationships, and the volume of goods and services exchanged. Among foragers and forager-horticulturalists, these patterns are most clearly manifest in the cooperative production and sharing of food (Gurven, 2004) and aid during illness and injury (Gurven et al., 2000, 2012; Sugiyama and Chacon, 2000). Hill and colleagues have argued that extensive exchange, facilitated by home-based resource sharing, is a universal pattern among hunter-gatherers (Hill, 2002; Hill et al., 2009). Among the Aché, for example, over 90 percent of the meat that hunters acquire is consumed by members of other nuclear families (Kaplan and Hill, 1985).
Anthropologists, economists, and biologists have not reached consensus regarding why exchange is so pervasive in human groups. Studies from human foragers have suggested that extensive food sharing within and among families relates to the specific feeding niche that foragers occupy. The foods that foragers exploit tend to be large, patchily distributed plant and animal species, whose availability can be clumped either in time or in space. The large package size (i.e., amount of food obtained at one time) and patchiness of foods is associated with large short-term fluctuations in food acquisition, with little or no food being acquired on many days followed by bounties on others. For example, the likelihoods of returning empty handed from a hunting attempt among Aché, Hadza, Hiwi, and Tsimane hunters are 96 percent, 65 percent, 45 percent, and 40 percent, respectively (Hill and Hurtado, 2009; Gurven et al., 2012). Shortfalls of no meat for a week or more would occur frequently for a single hunter and his family under these conditions; at the same time, a single family would be unable to consume a large bounty of meat before it spoiled.
Two different theories have been advanced to explain the extent of food sharing in hunting and gathering societies, both of which emphasize the importance of variability in food supply. According to one view, reciprocity-based risk reduction drives food transfers; according to the other, food transfers are the result of tolerated theft. The “reciprocity and risk reduction” view proposes that where foods are unable to be preserved or stored, the cycle of bounties and shortfalls can be buffered by reciprocal
food sharing (Kaplan and Hill, 1985; Winterhalder, 1986; Hames, 1990; Gurven, 2004). According to the theory, the challenge posed by intertemporal variability in large-package food availability was addressed culturally with an “insurance” mechanism: successful foragers of high-variability resources share food with unsuccessful foragers with the expectation of a reciprocal flow in the other direction when the situation is reversed. By smoothing variability in intake rates, foragers maximized the nutritional benefit from large food packages. The “tolerated theft” hypothesis proposes that large food packages create variation among individuals in their available food, with the “lucky” individuals possessing larger and the unlucky ones smaller quantities or none (Blurton Jones, 1984). Possessors tolerate theft from their supply by individuals with less, because the cost to the possessor of defending the surplus exceeds its value, and the value of the food to the hungry individual makes fighting for it worthwhile. Hence, exchange is not reciprocal, but based on momentary variation in who is hungry and who has abundance.
For reciprocity-based risk reduction to explain food transfers, it is necessary to show that producers can direct food transfers to preferred individuals, and those transfers are directed to those who have given previously. The bi-directionality (or “contingency”) of sharing has been evaluated by examining the extent to which the amounts given from one individual or family to another are correlated with amounts received in return, controlling for other factors such as kinship and spatial proximity. Positive contingency has been found in observational studies from the reservation Aché, Aka, Hadza, Hiwi, Meriam, Mikea, Miskito, Shuar, Tsimane, and Yanomamö (see meta-analysis of Jaeggi and Gurven, 2013).
Another way to examine the extent to which food transfers in hunting and gathering groups are governed by individually based social relationships in response to risk, is to examine how much food is given to different food-sharing partners as a function of the size of the food package. Figure 8-5 shows how small and large packages were handled by Aché foragers living at settlement after contact. The figure plots the percent of food items given to other families in rank order based on the total frequency of sharing to each family over time. This plot shows that the likelihood of giving depends on both the size of the game animal and rank order. Most-preferred partners (rank 1) received shares close to 50 percent of the time for both large and small game. However, less-preferred partners received shares less often for small game than for large game. For example, partners of rank 8 received a share only 5 percent of the time for small game, but close to 20 percent of the time for large game. This pattern would make sense if the size of the risk-sharing pool was strategically adjusted to package size, and if transfers were based on individually negotiated relationships (Kaplan and Gurven, 2005).
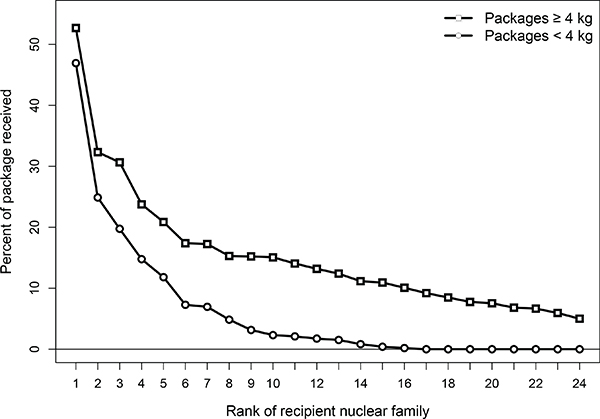
FIGURE 8-5 Aché food sharing by package size.
NOTE: The horizontal axis indicates the rank of recipient nuclear families from highest (rank 1) to lowest (rank 24) amount shared. The vertical axis indicates the mean percent of food items shared with each family. Large foods with more variable return rates are shared with a broader network of social partners than smaller, less variable foods.
SOURCE: Kaplan and Gurven (2005).
Returns to scale in the collective production of food also motivate the formation of reciprocal cooperative relationships and food sharing among human foragers. Many game species are effectively hunted through the cooperation of multiple hunters, which can allow capture of large game—such as giraffes or whales—or achieve multiple kills of animals that move in herds or groups—such as monkeys, coatis, peccaries, or larger ungulates. Among the Aché, approximately 50 percent of all game acquired is derived from cooperatively pursued species, with hunters often signalling each other upon encountering prey in order to recruit help (Hill, 2002). Among many pygmy groups, the majority of game is captured through cooperative game drives; similar drives have been described in western North America. Among the Lamalera in Indonesia, whales are hunted by large, well-organized pursuit groups in boats, and food shares are given to participants and other families according to well-prescribed rules (Alvard and Nolin,
2002). Cooperative whale hunting with concommitant food sharing has also been well-documented among the Inuit (Balikci, 1989; Smith, 1991).
There are two important implications of extensive food sharing and other forms of bilateral cooperation for health and well-being among foragers. First, sharing has important effects in reducing risks of mortality throughout life. Among the Tsimane, for example, food transfers and other forms of assistance are given to orphaned children (who are at risk of increased mortality), to women who have lost a spouse, to sick individuals until they recover, and following loss of food or possessions (Gurven et al., 2012). Sugiyama and Chacon’s study of Yora and Shiwiar forager-horticulturalists showed that illnesses and injuries prevented men from foraging on approximately 10 percent of person days. They found that not only would the sick or wounded individual and his family suffer from the reduced protein intake, but so too would others in his sharing network. On the basis of those results, they argued that foragers must maintain large sharing networks to buffer the risks of illness and injury (Sugiyama and Chacon, 2000).
Weissner’s analysis of Ju/’hoansi (!Kung) sharing networks came to similar conclusions, but on the basis of environmental risks. She found that Ju/’hoansi maintain sharing partners in a system called xharo, in which men and women commit to obligations of mutual aid and gift giving. She found that on average, Ju/’hoansi adults had 16 partners, with a range from 2 to 42 partners. Individual partner sets tended to include individuals of both sexes of varying ages, and with a variety of skills. Approximately 18 percent of partners resided in ego’s camp, 24 percent in neighboring camps 1-15 km away, 25 percent in camps 16-50 km away, and 33 percent in camps 51-200 km away. One advantage of maintaining partners in distant camps was that in times of local food or water shortage, xharo partners could be visited in distant places where conditions were better. She also found strong associations between foraging efficiency, food transfers, possessions, and reproductive success: Better hunters who transfered more food had 80 percent more xharo partners, received more material possessions though xharo, and had an average of 50 percent more children (Wiessner, 1982, 2002).
A second potential implication of food sharing, as a response to either risk or the gains from cooperative production, is that it may limit the scope for dominance-based social relationships and the formation of social hierarchies. In many social species, dominance is used to gain exclusive rights to resources. Among female primates, for example, rights to favored territories or foraging locations for themselves and their offspring are often subjects of dominance interactions (Pusey et al., 2008). Among foraging groups, dominance hierarchies and within-group disputes over access to resource patches are not often reported (Boehm, 2009; there are exceptions to be
discussed below). Among foragers, where the gains from cooperation are a major determinant of well-being, overt dominance behavior can bring the risk of losing valuable cooperative partners. It is likely to be difficult to coerce people into working and giving up shares of their food through dominance, particularly when resources are not concentrated and predictable. High mobility between groups, and frequent group fissioning and fusing, are commonly reported for foragers (e.g., Hill and Hurtado, 1996), which may also limit the scope for dominance. In the light of these factors, it appears that “egalitarianism” may emerge in part from the formation of voluntary partnerships, based on mutual support and joint production, but may not imply equal outcomes for all.
One final element may tie these threads together. The greater the importance of male foraging effort to the health and well-being of children, the less will be the gains to men from dominance interactions over mating opportunities. If, at equilibrium, an average man can only support all the children of one woman, the gains from dominance over other men may be small. The gains from risk reduction, cooperative acquisition, and male parental investment may thus combine to generate the relatively egalitarian patterns of interaction that seem to characterize so many foraging groups and that differ markedly from most other primate species.
VARIATION AMONG FORAGERS
To this point, we have discussed general trends in the social relationships of societies that depend on foraging for basic subsistence, and have attempted to describe what we refer to as the modal pattern. In this section, we ask whether the theoretical principles discussed above can explain variation (a) within the modal pattern and (b) away from the modal pattern.
With respect to intergenerational transfers, our discussion emphasized the implications of skills-based foraging for age-specific food production and, in turn, kin-based downward resource transfers. The logic presented above would imply that ecological variation in the skill-demands of the foods exploited, or in age-specific productivity profiles, would be associated with changes in transfers. There are a suite of ethnographic observations suggesting that variations within the modal pattern may be explained by such variation. Seasonal variation in fruit availability among the Aché, for example, is associated with the proportion of children’s diet that they can provide for themselves. During fruit seasons, food production for children under age 14 increases 5-fold, allowing them to produce more than half of the calories they consume (Kaplan, 1997). Similarly, intergenerational transfers to Hadza children appear to be smaller than to Ju/’hoansi children, because the former acquire more fruit. Part of this difference in turn has been attributed to the greater safety in Hadza environments for trav-
eling to and from food patches (Blurton Jones et al., 1994). Among the Mikea of Madagascar, many game animals are nocturnal and may be easily captured, even by children, making the children less dependent on parents (Tucker and Young, 2005). In spite of this variation within the modal pattern, we know of no variations away from the modal pattern of downward intergenerational transfers among foragers.
With respect to male parental investment and monogamy, there are fewer quantitative data to assess ecological trends. The expectations derived from the discussion above are that increased polygyny would occur for one of three reasons. One reason would be decreased gains from male parental investment, motivating women to choose males more on the basis of genetic quality, and motivating men to invest more in mating competition. The second reason is that, under conditions of high interpersonal violence, male protection may become a more critical input into wife and offspring success. Where a husband’s ability to protect has a direct impact on the well-being of his immediate family, and protection can be provided as an “umbrella” benefit (i.e., a common or nonrival good) to wives and offspring, female choice may shift, with greater preferences for males with greater ability to provide security (and less weight placed on more strictly rival goods, such as time and energy; Bowles and Borgerhoff Mulder, 2012). This could result in greater levels of polygyny because one man might be able to offer protection to several wives and their offspring. The third reason would be increased variaton among males in the parental investment they have to offer, leading “wealthier” (or otherwise higher-quality) males to have multiple wives, and others to have none.
Very high rates of polygyny have been reported for some Australian aboriginal groups and for some native North Americans living on the plains. The causes of high rates of polygyny in aboriginal Australia are not well understood, but Hart et al. (1960) suggested that women in the northern territories could be largely self-sufficient, again because hunting nocturnal animals was relatively easy. In the plains of North America, the adoption of the horse seems to be responsible for major changes in the economy, with large-scale implications for the mating system. Small foraging bands were converted into large raiding parties, stealing herds of horses and women at the same time. Wealthy men with large herds could also obtain additional wives through brideprice. In essence, they became partial pastoralists, with many convergences in social organization (Steward, 1938; Hämäläinen, 2009).
With respect to reciprocity and egalitarianism, there is well-documented variation in patterns of food transfer, both within and among groups. A robust predictor of the amount of food transferred in terms of sharing breadth (the number of partners receiving shares) is the size of food packages and the associated variability in their delivery (see Figure 8-5 and
review in Gurven, 2004). In cases where very large game are customary, such as the Hadza, food transfers often involved members from more than one camp or group (Hawkes et al., 2001). Seasonal variation in resource availability and patchiness can also have dramatic effects on this aspect of social organization. For example, Steward (1938) described that in the great basin of North America, people lived in largely self-sufficient, small nuclear-family clusters during the winter when resources were scarce and congregated in large groups, participating in communal game drives during the summer when resources permitted. Band-wide, fully communistic food sharing has also been observed, as in the case of the Aché and Yora during mobile foraging trips (but not at permanent settlements: Kaplan and Hill, 1985; Hill and Kaplan, 1989). In those cases, producers do not appear to control to whom they give, higher producers give more than they receive, and larger families receive bigger shares (Kaplan and Hill, 1985; Kaplan et al., 1990). Band-wide sharing of meat appears to occur when groups are relatively small, and strategies that maximize group returns through cooperative efforts require some individuals to give up the chance of making a kill.
Movement away from the egalitarian modal pattern has been documented in several places, most notably on the west coast of North America. Large differences in status and restricted access to resource patches based on inheritance have been reported for several groups living in the Pacific Northwest and the coast of California (Ames, 1994). One suggestion for explaining this trend away from egalitarianism toward hierarchy concerns the nature of variability in resource availability. Reciprocity is an effective strategy when variability across individuals is unsynchronized in time. When there is predictable variation in resource availability across space with reasonable intertemporal stability (e.g., salmon runs and acorn groves), there can be gains to investment in dominance in order to gain preferential access to rich resource patches (Dyson-Hudson and Smith, 1978; Kelly, 1995). Such differential access could be converted into inherited differences in wealth, and number of wives (Borgerhoff Mulder et al., 2009; Smith et al., 2010). One can note that the modern distribution of foraging groups is unlikely to encompass the full range of variation in the extent of competition for durable territorial resources possible in the pre-Neolithic past, and that more territorially focused foragers may have clustered in predictably rich areas (e.g., along coasts and rivers), with higher levels of social hierarchy and inequality.
Until now, we have discussed both full-time hunter-gatherers and forager-horticulturalists, who practice shifting cultivation, as conforming largely to the modal pattern. However, the inclusion of cultivation (i.e., farming) in the subsistence regime does seem to have some important downstream effects. For one, farming appears to have complex effects on intergenerational transfers. Among the Tsimane, men shift their effort from
hunting to farming as they age, reportedly because they can no longer hunt effectively (which is consistent with data on age-related decreases in hunting return rates: Hooper, 2011). This may suggest that farming allowed greater contributions of individuals in their 60s and 70s to younger individuals. However, there are farming tasks, such as harvesting, that children can do well, leading to increased productivity of children and an earlier age of nutritional independence (Kramer, 2005). Where farming reduces the importance of hunted foods in the diet, or male labor more generally, it may result in less male parental investment and more polygyny. The largest shifter, however, within farming groups is whether they own and defend territories. It is with territorial ownership that humans shift to forms of social organization outside the scope of this paper. The patterns discussed for “territory owning” hunter-gatherers in North America provide some insights into those transformations.
DISCUSSION AND CONCLUSIONS
The two central messages of this paper are, first, that human demography can only be well understood in light of human social structure, and vice versa; and second, that both of these domains have been conditioned by the ecologies inhabited by hunter-gatherers in the remote evolutionary past. In the preceding sections, we have drawn on a growing body of quantitative ethnographic research to discuss the evolutionary interdependencies among (1) longevity, menopause, and intergenerational transfers; (2) the coupling of male and female reproductive careers, biparental investment, and cooperative pair-bonds; and (3) survival and cooperative social insurance.
This synthesis may afford new insights into both human evolution and contemporary human behavior. Very little is known about the temporal sequence for the evolution of those features of social life that are close to universal among extant foragers. Given the isolation of peoples in the Americas for close to 15,000 years and in Australia for more than 30,000 years, it would appear that such traits are likely to have been present in the first anatomically modern humans. It appears that initial brain expansion and greater emphasis on meat eating occurred with the emergence of the genus Homo some 1.8-2 million years ago. Yet much remains to be learned about the co-evolution of these biosocial traits. Did they slowly ratchet each other, or were some traits in place long before others? The answers to these questions are likely to be found in the future with some combination of paleogenetics, isotype analysis, and other methods yet to be discovered. The theoretical principles discussed above can serve to guide such research.
The insights into modern behavior are likely to derive from two avenues. First, we expect there to be much “carry-over” or inertia from the past. While there is evidence of genetic changes since the advent of agriculture
and animal husbandry—such as with respect to lactose tolerance (Durham, 1991)—there seems to be strong canalization of human development with respect to brain development, growth, maturation, and aging. There are secular trends, but variation is limited: Nowhere are people mature at age 5 or living to age 200. Lee (2007) has argued that downward transfers within families are preserved across societal transitions, and the truly novel development is upward transfers, not within families, but through government institutions, such as Social Security and Medicare.
Monogamous marriage is now prescribed by law in most of the world, but to what extent do those cultural institutions have long evolutionary roots? The endocrinological mechanisms underlying a father’s attachment to his children (Gray and Campbell, 2009; Gettler et al., 2011) are likely the product of millenia of selection under past conditions. At the same time, patterns of opportunistic desertion and disinvestment by males indicate subtantial flexibility of strategies and adjustment to local socioecological context (Muller et al., 2009; Stieglitz et al., this volume). Similarly, there is growing evidence that social networks affect virtually every aspect of life and well-being in modern societies. To what extent are those patterns relics of the social past? These are questions that future research can address.
The second avenue of insight into modern human behavior may stem from the theoretical principles that appear to order the data collected among foraging societies. The gains from investments in skill and other factors affecting age profiles of productivity, such as health in old age, may help explain patterns of intergenerational transfers and parental investment that change over time and depend on socioeconomic status. The outcomes associated with biparental care compared with uniparental care—which vary across social, economic, and institutional contexts—may help explain the distribution and duration of marital/cohabiting and nonmarital/noncohabiting unions. Gains from social risk sharing and joint production may also provide insights into the nature of social interactions among non-kin, and into effort put into social dominance both within and among nations. There is still much to be learned regarding the legacy of the past.
REFERENCES
Alberts, S.C., Altmann, J., Brockman, D.K., Cords, M., Fedigan, L.M., Pusey, A., Stoinski, T.S., Strier, K.B., Morris, W.F., and Bronikowski, A.M. (2013). Reproductive aging patterns in primates reveal that humans are distinct. Proceedings of the National Academy of Sciences of the United States of America, 110(33), 13440-13445.
Alvard, M.S., and Nolin, D.A. (2002). Rousseau’s whale hunt? Coordination among big-game hunters. Current Anthropology, 43(4), 533-559.
Ames, K.M. (1994). The Northwest Coast: Complex hunter-gatherers, ecology, and social evolution. Annual Review of Anthropology, 23(1), 209-229.
Balikci, A. (1989). The Netsilik Eskimo. Prospect Heights, IL: Waveland Press, Inc.
Becker, G.S. (1973). A theory of marriage: Part I. The Journal of Political Economy, 81(4), 813-846.
Bennett, P.M., and Owens, I.P.F. (2002). Evolutionary Ecology of Birds. New York: Oxford University Press.
Binford, L.R. (2001). Constructing Frames of Reference. Oakland: University of California Press.
Blurton Jones, N.G. (1984). A selfish origin for human food sharing: Tolerated theft. Ethology and Sociobiology, 5(1), 1-3.
Blurton Jones, N.G., Smith, L.C., O’Connell, J.F., Hawkes, K., and Kamuzora, C.L. (1992). Demography of the Hadza, an increasing and high density population of Savanna foragers. American Journal of Physical Anthropology, 89(2), 159-181.
Blurton Jones, N.G., Hawkes, K., and Draper, P. (1994). Foraging returns of !Kung adults and children: Why didn’t !Kung children forage? Journal of Anthropological Research, 50(3), 217-248.
Boehm, C. (2009). Hierarchy in the Forest. Cambridge, MA: Harvard University Press.
Bogin, B.A., and Smith, B.H. (1996). Evolution of the human life cycle. American Journal of Human Biology, 8(6), 703-716.
Borgerhoff Mulder, M., Bowles, S., Hertz, T., Bell, A., Beise, J., Clark, G., Fazzio, I., Gurven, M., et al. (2009). Intergenerational wealth transmission and the dynamics of inequality in small-scale societies. Science, 326(5953), 682-688.
Bowles, S., and Borgerhoff Mulder, M. (2012). The Nature of Wealth, Wealth Inequality, and the Polygyny Paradox. Polygyny Paradox Project. Santa Fe, NM: Santa Fe Institute.
Clutton-Brock, T.H. (1991). The Evolution of Parental Care. Princeton, NJ: Princeton University Press.
Clutton-Brock, T.H., and Isvaran, K. (2007). Sex differences in ageing in natural populations of vertebrates. Proceedings of the Royal Society B: Biological Sciences, 274(1629), 3097-3104.
Durham, W.H. (1991). Coevolution: Genes, Culture, and Human Diversity. Redwood City, CA: Stanford University Press.
Dyson-Hudson, R., and Smith, E.A. (1978). Human territoriality: An ecological reassessment. American Anthropologist, 80(1), 21-41.
Foote, A.D. (2008). Mortality rate acceleration and post-reproductive lifespan in matrilineal whale species. Biology Letters, 4(2), 189-191.
Gettler, L.T., McDade, T.W., Feranil, A.B., and Kuzawa, C.W. (2011). Longitudinal evidence that fatherhood decreases testosterone in human males. Proceedings of the National Academy of Sciences of the United States of America, 108(39), 16194-16199.
Grafen, A., and Sibly, R. (1978). A model of mate desertion. Animal Behaviour, 26, 645-652.
Gray, P.B., and Campbell, B.C. (2009). Human male testosterone, pair bonding and fatherhood. In P.T. Ellison and P.B. Gray (Eds.), Endocrinology of Social Relationships (pp. 270-293). Cambridge, MA: Harvard University Press.
Gross, M.R., and R.C. Sargent. (1985). The evolution of male and female parental care in fishes. American Zoologist, 25(3), 807-822.
Gurven, M. (2004). To give and to give not: The behavioral ecology of human food transfers. Behavioral and Brain Sciences, 27(04), 543-559.
Gurven, M., and Kaplan, H. (2006). Determinants of time allocation to production across the lifespan among the Machiguenga and Piro indians of Peru. Human Nature, 17(1), 1-49.
Gurven, M., and Kaplan, H. (2007). Longevity among hunter-gatherers: A cross-cultural examination. Population and Development Review, 33(2), 321-365.
Gurven, M., and Walker, R. (2006). Energetic demand of multiple dependents and the evolution of slow human growth. Proceedings of the Royal Society B: Biological Sciences, 273(1588), 835-841.
Gurven, M., Allen-Arave, W., Hill, K., and Hurtado, A.M. (2000). “It’s a wonderful life”: Signaling generosity among the Ache of Paraguay. Evolution and Human Behavior, 21(4), 263-282.
Gurven, M., Kaplan, H., and Gutierrez, M. (2006). How long does it take to become a proficient hunter? Implications for the evolution of extended development and long life span. Journal of Human Evolution, 51(5), 454-470.
Gurven, M., Stieglitz, J., Hooper, P.L., Gomes, C., and Kaplan, H. (2012). From the womb to the tomb: The role of transfers in shaping the evolved human life history. Experimental Gerontology, 47(10), 807-813.
Hämäläinen, P. (2009). The Comanche Empire. New Haven, CT: Yale University Press.
Hames, R. (1990). Sharing among the Yanomamö: Part I, The effects of risk. In E. Cashdan (Ed.), Risk and Uncertainty in Tribal and Peasant Economies (pp. 89-106). Boulder, CO: Westview Press.
Hart, C.W.M., Pilling, A.R., and Goodale, J.C. (1960). The Tiwi of North Australia. New York: Holt, Rinehart and Winston.
Hawkes, K., O’Connell, J.F., and Blurton Jones, N.G. (1989). Hardworking Hadza grandmothers. Comparative Socioecology, 341-366.
Hawkes, K., O’Connell, J.F., and Blurton Jones, N.G. (1997). Hadza women’s time allocation, offspring provisioning, and the evolution of long postmenopausal life spans. Current Anthropology, 38(4), 551-577.
Hawkes, K., O’Connell, J.F., Blurton Jones, N.G., Alvarez, H., and Charnov, E.L. (1998). Grandmothering, menopause, and the evolution of human life histories. Proceedings of the National Academy of Sciences of the United States of America, 95(3), 1336-1339.
Hawkes, K., O’Connell, J.F., and Blurton Jones, N.G. (2001). Hadza meat sharing. Evolution and Human Behavior, 22(2), 113-142.
Headland, T.N. (1986). Why Foragers Do Not Become Farmers: A Historical Study of a Changing Ecosystem and Its Effect on a Negrito Hunter-Gatherer Group in the Philippines. Ph.D. Thesis. Honolulu: University of Hawaii.
Headland, T.N., and Reid, L.A. (1991). Holocene foragers and interethnic trade: A critique of the myth of isolated independent hunter-gatherers. In S.A. Gregg (Ed.), Between Bands and States (pp. 333-340). Carbondale: Southern Illinois University.
Hewlett, B.S. (1992). Husband-wife reciprocity and the father-infant relationship among Aka pygmies. In B.S. Hewlett (Ed.), Father-Child Relations: Cultural and Biosocial Contexts (pp. 53-176). New Brunswick, NJ: Transaction Publishers.
Hewlett, B.S., and Macfarlan, S.J. (2010). Fathers’ roles in hunter-gatherer and other small-scale cultures. In M.E. Lamb (Ed.), The Role of the Father in Child Development (pp. 413-434). Hoboken, NJ: John Wiley & Sons, Inc.
Hill, K. (2002). Altruistic cooperation during foraging by the Ache, and the evolved human predisposition to cooperate. Human Nature, 13(1), 105-128.
Hill, K., and Hurtado, A.M. (1996). Ache Life History. New Brunswick, NJ: Transaction Publishers.
Hill, K., and Hurtado, A.M. (2009). Cooperative breeding in South American hunter-gatherers. Proceedings of the Royal Society B: Biological Sciences, 276(1674), 3863-3870.
Hill, K., and Kaplan, H. (1989). Population description and dry season subsistence among the newly contacted Yora (Yaminahua) of Manu National Park, Peru. National Geographic Research, 5(3), 317-334.
Hill, K., Hurtado, A.M., and Walker, R.S. (2007). High adult mortality among Hiwi hunter-gatherers: Implications for human evolution. Journal of Human Evolution, 52, 443-454.
Hill, K., Barton, M., and Hurtado, A.M. (2009). The emergence of human uniqueness: Characters underlying behavioral modernity. Evolutionary Anthropology, 18(5), 187-200.
Hooper, P.L. (2011). The Structure of Energy Production and Redistribution among Tsimane´ Forager-Horticulturalists. Ph.D. Thesis. Albuquerque: University of New Mexico.
Hooper, P.L., Gurven, M., Winking, J., and Kaplan, H.S. (2014). Inclusive Fitness Explains Intergenerational Transfers in a Traditional Human Society. SFI Working Paper. Santa Fe, NM: Santa Fe Institute.
Hooper, P.L., Lancaster, J.B., Gurven, M., and Kaplan, H.S. (2015). Cooperation between the sexes. In M. Muller, R. Wrangham, and D. Pilbeam (Eds.), Chimpanzees and Human Evolution. Cambridge, MA: Harvard University Press.
Howell, N. (1979). Demography of the Dobe !Kung. New Brunswick, NJ: Transaction Publishers.
Howell, N. (2010). Life Histories of the Dobe !Kung. Oakland: University of California Press.
Hrdy, S.B. (2009). Mothers and Others. Cambridge, MA: Belknap Press of Harvard University Press.
Hurtado, A.M., Hawkes, K., Hill, K., and Kaplan, H. (1985). Female subsistence strategies among Ache hunter-gatherers of Eastern Paraguay. Human Ecology, 13(1), 1-28.
Hurtado, A.M., Hill, K., Hurtado, I., and Kaplan, H. (1992). Trade-offs between female food acquisition and child care among Hiwi and Ache foragers. Human Nature, 3(3), 185-216.
Jaeggi, A.V., and Gurven, M. (2013). Natural cooperators: Food sharing in humans and other primates. Evolutionary Anthropology, 22(4), 186-195.
Jaeggi, A.V., and Van Schaik, C.P. (2011). The evolution of food sharing in primates. Behavioral Ecology and Sociobiology, 65(11), 2125-2140.
Jaeggi, A.V., Van Noordwijk, M.A., and Van Schaik, C.P. (2008). Begging for information: Mother-offspring food sharing among wild Bornean orangutans. American Journal of Primatology, 70(6), 533-541.
Kaplan, H. (1994). Evolutionary and wealth flows theories of fertility: Empirical tests and new models. Population and Development Review, 753-791.
Kaplan, H. (1997). The evolution of the human life course. In K.W. Wachter and C.E. Finch (Eds.), Between Zeus and the Salmon: The Biodemography of Longevity (pp. 175-211). Committee on Population, Commission on Behavioral and Social Sciences and Education. Washington, DC: National Academy Press.
Kaplan, H., and Gurven, M. (2005). The natural history of human food sharing and cooperation: A review and a new multi-individual approach to the negotiation of norms. In H. Gintis, S. Bowles, R. Boyd, and E. Fehr (Eds.), Moral Sentiments and Material Interests: The Foundations of Cooperation in Economic Life (pp. 75-113). Cambridge, MA: The MIT Press.
Kaplan, H., and Hill, K. (1985). Food sharing among Ache foragers: Tests of explanatory hypotheses. Current Anthropology, 223-246.
Kaplan, H., and Robson, A.J. (2002). The emergence of humans: The coevolution of intelligence and longevity with intergenerational transfers. Proceedings of the National Academy of Sciences of the United States of America, 99(15), 10221-10226.
Kaplan, H., Hill, K., and Hurtado, A.M. (1990). Risk, foraging and food sharing among the Ache. In E. Cashdan (Ed.), Risk and Uncertainty in Tribal and Peasant Economies (pp. 107-144). Boulder, CO: Westview Press.
Kaplan, H., Hill, K., Lancaster, J., and Hurtado, A.M. (2000). A theory of human life history evolution: Diet, intelligence, and longevity. Evolutionary Anthropology, 9(4), 156-185.
Kaplan, H., Hill, K., Hurtado, A.M., and Lancaster, J.B. (2001). The embodied capital theory of human evolution. In P.T. Ellison (Ed.), Reproductive Ecology and Human Evolution (pp. 293-318). Hawthorne, NY: Aldine de Gruyter.
Kaplan, H., Gurven, M., Winking, J., Hooper, P.L., and Stieglitz, J. (2010). Learning, menopause, and the human adaptive complex. Annals of the New York Academy of Sciences, 1204(1), 30-42.
Kelly, R.L. (1995). The Foraging Spectrum. Washington, DC: Smithsonian Institution Press.
Kidd, M.R., Duftner, N., Koblmüller, S.., Sturmbauer, C., and Hofmann, H.A. (2012). Repeated parallel evolution of parental care strategies within Xenotilapia, a genus of cichlid fishes from Lake Tanganyika. PLOS ONE, 7(2), e31236.
Kokko, H., and Jennions, M.D. (2008). Parental investment, sexual selection and sex ratios. Journal of Evolutionary Biology, 21(4), 919-948.
Kokko, H., and Johnstone, R.A. (2002). Why is mutual mate choice not the norm? Operational sex ratios, sex roles and the evolution of sexually dimorphic and monomorphic signalling. Philosophical Transactions of the Royal Society B: Biological Sciences, 357(1419), 319-330.
Kramer, K. (2005). Maya Children. Cambridge, MA: Harvard University Press.
Lancaster, J.B., and King, B.J. (1985). An evolutionary perspective on menopause. In J. Brown and V. Kerns (Eds.), In Her Prime: A New View of Middle Aged Women (pp. 13-20). Westport, CT: Bergin & Garvey.
Lancaster, J.B., and Lancaster, C.S. (1983). Parental investment: The hominid adaptation. In D.J. Ortner (Ed.), How Humans Adapt: A Biocultural Odyssey (pp. 33-56). Washington, DC: Smithsonian Institution Press.
Lazarus, J. (1990). The logic of mate desertion. Animal Behaviour, 39(4), 672-684.
Lee, R.D. (2007). Demographic change, welfare, and intergenerational transfers: A global overview. In J. Véron, S. Pennec, and J. Légaré (Eds.), Ages, Generations and the Social Contract (pp. 17-43). Dordrecht, The Netherlands: Springer.
Marlowe, F.W. (2003). A critical period for provisioning by Hadza men: Implications for pair bonding. Evolution and Human Behavior, 24(3), 217-229.
Marlowe, F.W. (2007). Hunting and gathering the human sexual division of foraging labor. Cross-Cultural Research, 41(2), 170-195.
Marlowe, F.W. (2010). The Hadza. Oakland: University of California Press.
Maynard Smith, J. (1977). Parental investment: A prospective analysis. Animal Behaviour, 25, 1-9.
Melancon, T.F. (1982). Marriage and Reproduction Among the Yanomamö Indians of Venezuela. Ph.D. Thesis. State College: Pennsylvania State University.
Minge-Klevana, W. (1980). Does labor time decrease with industrialization? A survey of time-allocation studies. Current Anthropology, 279-298.
Muller, M.N., Marlowe, F.W., Bugumba, R., and Ellison, P.T. (2009). Testosterone and paternal care in East African foragers and pastoralists. Proceedings of the Royal Society B: Biological Sciences, 276(1655), 347-354.
Murdock, G.P., and Provost, C. (1973). Factors in the division of labor by sex: A cross-cultural analysis. Ethnology, 203-225.
Nishida, T., and Turner, L.A. (1996). Food transfer between mother and infant chimpanzees of the Mahale Mountains National Park, Tanzania. International Journal of Primatology, 17(6), 947-968.
Patton, J.Q. (2000). Reciprocal altruism and warfare: A case from the Ecuadorian Amazon. In L. Cronk, N. Chagnon, and W. Irons (Eds.), Adaptation and Human Behavior: An Anthropological Perspective (pp. 417-436). Hawthorne, NY: Aldine de Gruyter.
Price, E.C., and Feistner, A.T.C. (1993). Food sharing in lion tamarins: Tests of three hypotheses. American Journal of Primatology, 31(3), 211-221.
Pusey, A., Murray, C., Wallauer, W., Wilson, M., Wroblewski, E., and Goodall, J. (2008). Severe aggression among female Pan troglodytes schweinfurthii at Gombe National Park, Tanzania. International Journal of Primatology, 29, 949-973.
Rendell, L., and Whitehead, H. (2001). Culture in whales and dolphins. Behavioral and Brain Sciences, 24(02), 309-324.
Robson, A.J., and Kaplan, H.S. (2003). The evolution of human life expectancy and intelligence in hunter-gatherer economies. American Economic Review, 93(1), 150-169.
Silk, J.B. (1978). Patterns of food sharing among mother and infant chimpanzees at Gombe National Park, Tanzania. Folia Primatologica, 29(2), 129-141.
Smith, E.A. (1991). Inujjuamiunt Foraging Strategies: Evolutionary Ecology of an Arctic Hunting Economy. New Brunswick, NJ: Transaction Publishers.
Smith, E.A., Borgerhoff Mulder, M., Bowles, S., Gurven, M., Hertz, T., and Shenk, M.K. (2010). Production systems, inheritance, and inequality in premodern societies. Current Anthropology, 51(1), 85-94.
Steward, J.H. (1938). Basin-plateau Aboriginal Sociopolitical Groups. Washington, DC: U.S. Government Printing Office.
Sugiyama, L.S., and Chacon, R. (2000). Effects of illness and injury on foraging among the Yora and Shiwiar: Pathology risk as adaptive problem. In L. Cronk, N. Chagnon, and W. Irons (Eds.), Adaptation and Human Behavior: An Anthropological Perspective (pp. 371-395). Hawthorne, NY: Aldine de Gruyter.
Sussman, R.W., and Cloninger, C.R. (Eds.). (2011). Origins of Altruism and Cooperation. New York: Springer.
Tucker, B., and Young, A.G. (2005). Growing up Mikea. In M.E. Lamb and B.S. Hewlett (Eds.), Hunter-Gatherer Childhoods: Evolutionary, Developmental, and Cultural Perspectives (pp. 147-171). New Brunswick, NJ: Transaction Publishers.
Tuljapurkar, S.D., Puleston, C.O., and Gurven, M.D. (2007). Why men matter: Mating patterns drive evolution of human lifespan. PLOS ONE, 2(8), e785.
Walker, R., Hill, K., Kaplan, H., and McMillan, G. (2002). Age-dependency in hunting ability among the Ache of Eastern Paraguay. Journal of Human Evolution, 42(6), 639-657.
Wiessner, P. (1982). Beyond willow smoke and dogs’ tails: A comment on Binford’s analysis of hunter-gatherer settlement systems. American Antiquity, 171-178.
Wiessner, P. (2002). Hunting, healing, and hxaro exchange: A long-term perspective on !Kung (Ju/’hoansi) large-game hunting. Evolution and Human Behavior, 23(6), 407-436.
Williams, G.C. (1957). Pleiotropy, natural selection, and the evolution of senescence. Evolution, 11, 398-411.
Winking, J., Gurven, M., Kaplan, H., and Stieglitz, J. (2009). The goals of direct paternal care among a South Amerindian population. American Journal of Physical Anthropology, 139(3), 295-304.
Winterhalder, B. (1986). Diet choice, risk, and food sharing in a stochastic environment. Journal of Anthropological Archaeology, 5(4), 369-392.
Wood, B.M., and Marlowe, F.W. (2013). Household and kin provisioning by Hadza men. Human Nature, 24(3), 280-317.
Wrangham, R.W., Wilson, M.L., and Muller, M.N. (2006). Comparative rates of violence in chimpanzees and humans. Primates, 47(1), 14-26.




























