
5
Relation to Other Fields of Physics
For decades, biological physics was seen as sitting at the interface between physics and biology. As discussed in Part I, this location on the map of science does not do justice to the field: The physics of living systems now stands on its own as a branch of physics, rather than being a temporary alliance at the border between disciplines. The phenomena of life provide a continual supply of challenges to our fundamental understanding of physics. Responding to these challenges requires new concepts, new principles, new theoretical and experimental techniques, and new instruments. While biological physics has an independent existence, many of these new developments are deeply connected to progress in other fields, and this is part of the beauty of physics. This chapter surveys, briefly, some of the many points of contact between biological physics and the broader physics community, summarized even more briefly in Table 5.1.
In this chapter, as elsewhere in the report, there are subtleties in drawing boundaries. Part I of this report has defined biological physics as bringing the physicist’s style of inquiry to bear on the phenomena of life, asking the kinds of questions and searching for the kinds of answers that characterize our understanding of the inanimate world. But in this search for a physics of life, the community often has found questions and answers that have resonance with other areas of physics, or perhaps define a new subfield that overlaps with biological physics. Living systems also may provide a clear, even pedagogical example of concepts from other parts of physics, or crucial applications of new experimental methods. This chapter is devoted to this rich exchange between biological physics and the rest of physics. Part of the emergence of biological physics as a branch of physics is that this exchange goes in both
TABLE 5.1 Connections to Other Fields of Physics
| Other Physics Fields | Relationship to Biological Physics |
|---|---|
| Elementary particle physics | Source of experimental tools and theoretical ideas now used in biological physics; recipient of data analysis methods based on neural network ideas from the biological physics community. |
| Nuclear physics | One of the sources for post–World War II funding for interactions between physics and biology. |
| Atomic, molecular, and optical physics | Source for spectroscopic techniques and the evolution of microscopy; interaction led to Nobel Prizes in super-resolution images (Chemistry, 2014) and optical tweezers (Physics, 2018). |
| Statistical physics | Biological physics as a source of general statistical physics problems; living systems as decisive testing grounds for new theoretical ideas. |
| Soft condensed matter physics | Polymers, membranes, self-assembly, and other soft matter problems inspired by phenomena in the living world. |
directions, with the physics of living systems being both consumer and producer of ideas and methods whose primary impact is outside the field.
It might seem far-fetched to think that biological physics is connected to elementary particle physics. But the detectors that are used in today’s X-ray diffraction experiments on protein structure, for example, have their roots in particle physics experiments. Conversely, the machine learning methods that are used today in the analysis of raw data from particle physics have their roots in models of brain function that began in the biological physics community. On the theoretical side, ideas from quantum field theory are central to thinking about the collective behavior of flocks and swarms, for example, but this reflects the merging of field theory and statistical physics that occurred in the 1970s, in the wake of the renormalization group. The fact that one can understand aspects of a flock’s behavior, quantitatively, by thinking about broken symmetry and massless excitations (Chapter 3) is testimony to the unifying power of theoretical physics.
For many years after World War II, significant interactions between physics and biology were supported by the U.S. government as an adjunct to nuclear physics. This work included the production of rare isotopes that were used as tracers in explorations of many different living systems, and the study of the effects of radiation on organisms, which led to the discovery of basic DNA repair mechanisms. For the biological physics community this is largely a closed chapter, but it is fertile ground for historians of science.1
The emergence of biological physics in the modern form described in this report is approximately contemporary with the renaissance of atomic, molecular,
___________________
1 See, for example, A.N.H. Creager, 2014, Life Atomic: A History of Radioisotopes in Science and Medicine, University of Chicago Press, Chicago, IL.
and optical (AMO) physics, including observation of the first Bose-Einstein condensates. By then there had already been two or three decades of dramatic impacts of laser physics on the exploration of the living world. Notably, as fast pulse lasers were being developed, one of the first experimental targets for these instruments was the photosynthetic reaction center (Chapter 1). More generally, lasers made possible an enormous expansion of spectroscopic techniques, including many that are widely used in the study of living systems. Even more profound was the impact on microscopy. Lasers are essential to the widespread use of confocal microscopy, and high-powered pulsed lasers are essential for two-photon fluorescence microscopy, which allows high-resolution imaging deep into highly scattering media, such as the brain. Total internal reflection microscopy, nonlinear methods such as coherent anti-Stokes Raman microscopy and stimulated Raman scattering microscopy, fluorescence speckle microscopy, light sheet microscopy, structured illumination microscopy, and fluorescence recovery after photobleaching (FRAP) all have had substantial impact on biological physics, and the continued development of these methods by the biological physics community has had an impact on biology more broadly (Chapter 6).
Some of the most profound interactions between AMO and biological physics involved developing a deeper understanding of basic physical principles. Although it was known, in principle, that the “diffraction limit” was not really a limit on our ability to reconstruct details in images, it was a revolution to develop methods whereby such super-resolution images become routine (see Figure I.2). These methods, based on stochastic localization or stimulated emission depletion, were recognized by a Nobel Prize in 2014. Similarly, although it was known in the 19th century that beams of light can apply forces to objects, it would take until the late 20th century to understand the conditions under which these forces could trap neutral particles. Crucially, optical traps or tweezers can produce forces on the scale of those generated by single biological molecules, which opened a whole generation of new experiments in the biological physics community (Chapters 1 and 2); this development was recognized by a Nobel Prize in Physics in 2018 (see Box 1.3).
Single molecule manipulation, both with optical and with magnetic traps, made it possible to connect one of the classic problems of statistical mechanics with controlled, quantitative experiments. All textbooks on the subject consider the random chain polymer, which is a compelling illustration of entropic forces. The calculation typically imagines taking a single long molecule and pulling on it, and the theory generates predictions for the force or stiffness. With single molecule manipulation, one can do exactly this experiment, and one can use DNA synthesis to be sure that the polymer is precisely N units long. As shown in Figure 5.1, experiments are sufficiently accurate that one can resolve the difference between a full elastic theory of the polymer and the simplifying freely jointed chain model.
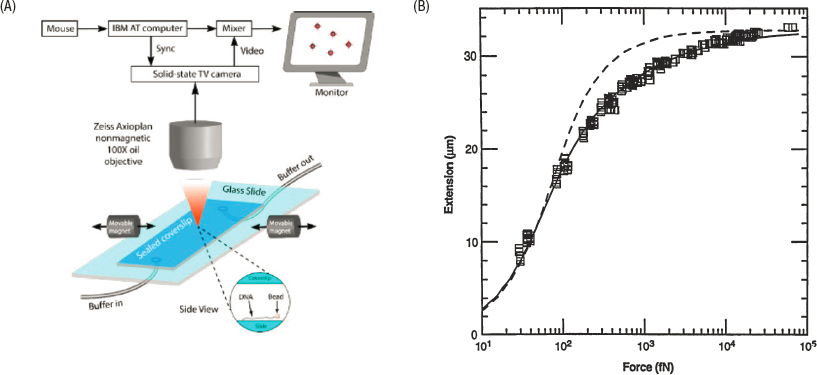
Polymers with N ∼100,000 units, as in Figure 5.1, are at the border between the molecules of AMO physics and the matter of condensed matter physics. Condensed matter physics is divided into “hard” and “soft,” where hard condensed matter is focused on problems where quantum mechanics is crucial, notably the behavior of electrons in solids. Soft condensed matter today encompasses many-body problems in the classical (non-quantum) limit. Many problems involving polymers, membranes, fluids, glasses, and other systems with complex landscapes are classical statistical physics problems, as with the polymer example in Figure 5.1. The soft matter community has substantial overlap with the biological physics community, and many soft matter problems are interesting in part because they capture some aspects of living systems in simpler and more easily controlled contexts.
As noted in Chapter 3, ideas from polymer physics also have been important in thinking about the structure and dynamics of chromosomes. These polymers are dense, and their configurations are shaped in part by active, non-equilibrium mechanisms. The nuclear pore complexes, which control transport in and out of cell nuclei, provide examples of polymer brushes, and it now is thought that different states of the brush can be identified with open and closed states of the pore, but
again it is essential that these systems are driven away from equilibrium, allowing active sorting of molecules between the cell nucleus and cytoplasm.
Early interest in another soft matter system, surfactant solutions, was motivated partly by oil recovery techniques and partly by cellular membranes composed of lipid bilayers. These systems often exhibit structure at mesoscopic scales, driven by competition between different forms of interactions, such as the hydrophilic and hydrophobic interactions in the case of surfactants or lipids. The existence of that structure and its sensitivity to thermal fluctuations or other perturbations lies at the core of soft matter research conducted in the 1980s and 1990s. There are echoes of this work in the current excitement surrounding phase transitions in real cell membranes and liquid-liquid phase separation in the cytoplasm (Chapter 3).
The complex phases that emerge in lipids and surfactants are examples of self-assembly. Although the structures are intricate, there is no blueprint. Self-assembly is an important meeting ground for the biological physics and soft matter physics communities. Living systems offer an astonishing array of structures that self-assemble, and using the term more loosely one can even think about structures that are actively constructed by communities of organisms, as in Figure 3.15. From the soft matter perspective, it is attractive to start with the very simplest examples and try to build up.
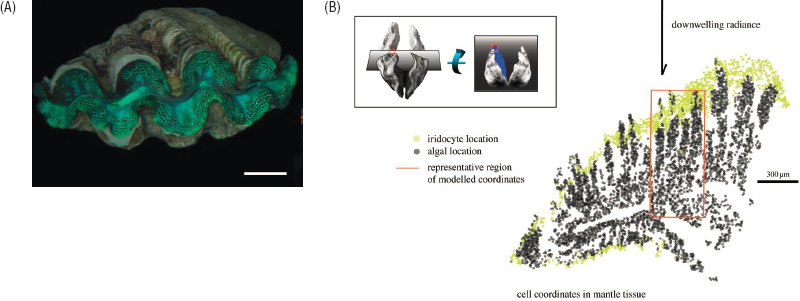
An example of self-assembly is provided by the lenses in the eyes of many aquatic animals, including cephalopods. Here the functional goal of self-assembly is to build a structure with desirable optical properties. Light passing through a curved lens is refracted more at the edges, distorting the image (spherical aberration). To avoid this, these eye lenses have an index of refraction that depends on the curvature. The squid lens is made of a single type of protein, but with subtle variations in amino acid sequence. These proteins have largely repulsive interactions, and the variations are in linker regions that bring the proteins into contact, allowing the formation of a gel. Variations in linker density drive gradients in the density of the gel and hence refractive index, avoiding aberration but also avoiding condensation into phases that would be opaque. Living systems make use of an astonishing variety of physical principles for building optical devices, in part because there are no intrinsically reflective parts such as metal surfaces. In insects, single photoreceptor cells can act as optical waveguides, pigment granules are transported through evanescent waves to act as attenuators (as with our pupil), stacks of cells and extracellular structures serve as interference-based reflectors, and more. In other examples, there is self-assembly of populations of organisms to perform crucial optical functions, as with the symbiosis between clams and algae in Figure 5.2, which also self-assemble.
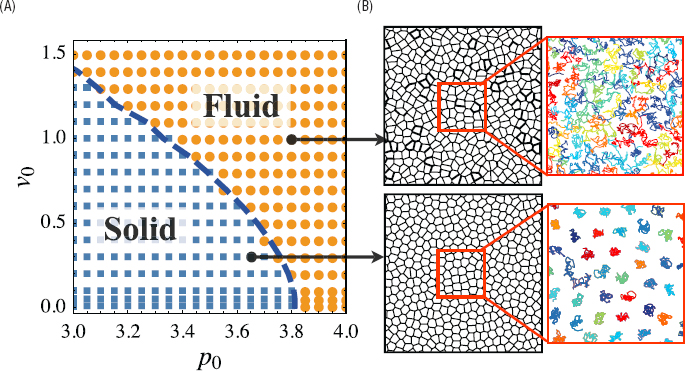
Phases and phase transitions are central to classical statistical mechanics, and a major theme of soft matter physics is to push these concepts beyond their original context of thermal equilibrium. A paradigmatic example is the jamming transition
in collections of randomly packed particles, which has the literal feel of a liquid/solid transition, but in the absence of a temperature or thermal motion. Epithelial tissues also undergo something like a liquid/solid transition, and recent work emphasizes that this rigidity transition, like jamming, is from a fluid to a rigid disordered state rather than an ordered one (see Figure 5.3).
The classical statistical physics of disordered systems connects with biological physics through many problems beyond cell movement. Understanding of glassiness in random heteropolymers provides a benchmark for thinking about protein folding, ultimately leading to the view that amino acid sequences must be selected by evolution to avoid the glass transition (Chapter 3). This example encourages thinking more generally about disordered systems in which the underlying randomness is shaped by some non-trivial physical constraints. Proteins do not just fold, they also exhibit allostery, in which the binding of a regulatory molecule to one site on a protein can influence the structure at very distant sites, for example changing the binding energy for a second type of molecule there. Theoretical approaches used to explore complex energy landscapes in disordered systems can be exploited to tune mechanical networks to display analogous allosteric behavior.
Theories of neural networks are a rich source of problems in statistical physics, so much so that one of the categories on the electronic archive of physics papers, arXiv.org, is “neural networks and disordered systems.” The relation of these statistical physics problems to real brains is discussed in Chapter 3, and application to the deep networks that are powering the current revolution in artificial intelligence is discussed in Chapter 7. Recent attempts to build analytic theories of structural glasses in high spatial dimension—models that might provide a mean-field approximation to real glasses—make strong connections to transitions that occur in neural network models, closing the circle.
Living systems are always away from equilibrium, and many crucial functions occur on a scale far from the thermodynamic limit, where the impact of single molecules can be felt clearly. As discussed in Part I of this report, the identification of these functions as physics problems, and understanding how they emerge, are central to biological physics. But the past two decades also have seen an explosion of activity in the statistical physics of small, non-equilibrium systems, more generally, often taking inspiration from the phenomena of life. The study of fluctuations in these systems has led to a number of remarkable results. These “fluctuation theorems” show, for example, that the distribution of microscopic response of
far-from-equilibrium systems contains equilibrium information, and this can be extracted by comparing processes and their time-reversed versions. Some of the first and still clearest tests of these ideas are in experiments with single biological molecules, adapting methods that were developed in the biological physics community, as for example in Figure 5.4. This stream of work also has emphasized that the equivalence of entropy as a measure of heat flow and entropy as a measure of available information is more than formal, and has physical consequences. Subsequent theoretical work has derived thermodynamic uncertainty relations, which for example connect the precision of molecular scale clocks to the rate of energy dissipation. There is a widespread sense in the community that all these ideas should feed back into our understanding of living systems, for example providing a new class of physical limits that can be compared to the performance of real molecular
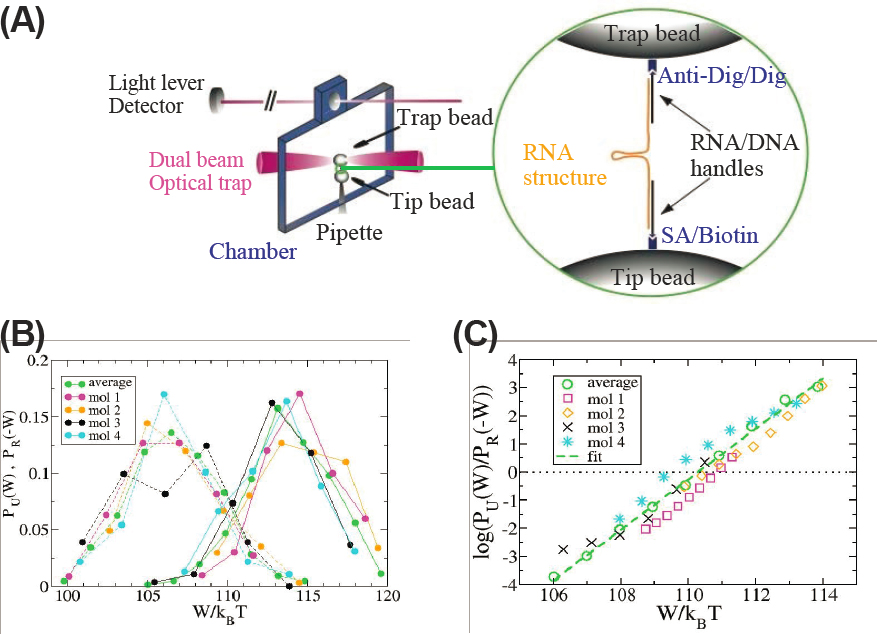
clocks such as the cell cycle or circadian rhythms, and more generally providing insight into the energetics of biochemical control.
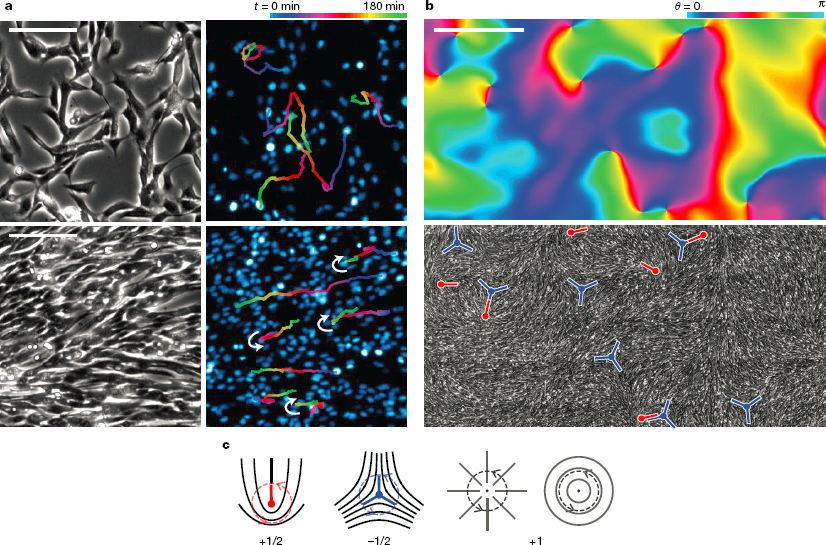
On a larger scale, physicists long wondered whether the agreement of birds in a flock to fly in the same direction could be seen as a transition to order, perhaps analogous to the alignment of spins in a magnet, but different because the “spins” (velocity vectors for the individual birds) are moving. As explained in Chapter 3, in the 1990s physicists began to study models that embody this intuition, and this launched the now lively field of active matter. The theory of active matter focuses on the derivation of effective theories for the generic behavior of these systems on length scales long compared with their component parts or the range of interactions. The renormalization group tells us that these behaviors should be independent of the short distance details. Importantly this means that one can observe related phenomena in flocks of birds and in schools of fish, in networks of protein filaments and motor molecules in single cells, and in populations of cells organizing themselves on a surface, as in Figure 5.5. In some cases, these predictions of universal behaviors are confirmed, while in other cases living systems have managed to circumvent generic expectations. This is generating a productive dialogue between the biological physics and active/soft matter communities, even driving the development of synthetic active systems.
Understanding animal movement depends on understanding the environment in which this movement happens. For organisms that swim or fly, this leads to potentially challenging problems in fluid mechanics, but these problems rest on a firm foundation. Many organisms, however, move on or through granular environments such as sand and soil. These materials flow, shift, and crack under stress, prototypes of the complex rheology that is a major theme in soft matter physics. Granular materials are collections of particles that interact via repulsive frictional contact forces; they remain solid below a yield stress, and flow frictionally above this stress. Diverse organisms, from ants to camels, contend with and utilize the properties of these soft materials. A newly hatched juvenile sea turtle, using its flippers to make its way to the ocean, must not drive the sand beyond its yield stress, and deadly sidewinder rattlesnakes climbing sandy desert dunes face the same problem. In contrast, slithering snakes and sand-swimming lizards create localized “fluids” and propel themselves within these. In granular systems, dissipation from frictional interactions dominates inertia, so that these quite macroscopic creatures face some of the same problems as bacteria, living at low Reynolds number (Chapter 1). In a very different regime of soft material behavior, a snail takes advantage of shear thinning in viscoelastic fluid as waves of muscle activity propagate along its foot. In all these examples, the physics of soft materials and the physics of animal behavior in relation to these materials are advancing together.
As emphasized above, a distinguishing feature of soft matter physics is that it is essentially classical rather than quantum mechanical. A related but distinct set
of questions is addressed by the nonlinear dynamics community. The problems of nonlinear dynamics have a venerable history, reaching back to the three-body problem in celestial mechanics. Many of the issues of stability and ergodicity, despite obvious physical consequences, became the province of pure mathematics for much of the 20th century, while fluid turbulence drifted out of physics into engineering. But there was a resurgence of interest from the physics community in the 1970s and 1980s, driven by a combination of theoretical and experimental developments. There has been a productive interplay between nonlinear dynamics and biological physics ever since.
Perhaps the most dramatic development in the resurgence of nonlinear dynamics as part of physics was the demonstration that there are a limited number of ways in which systems with small numbers of variables could make the transition from regular to irregular or chaotic motion. This work resulted in a complete analysis of these transitions using the renormalization group, and the verification of the theory in experiments on fluids and other systems. While the motivation for this work clearly came from the search for a more manageable version of the transition to turbulence, the low dimensional dynamical systems that were studied first came equally from fluid mechanics and from the dynamics of population growth. The tools that were built up to analyze low dimensional dynamical systems—bifurcations; fixed points and attractors; periodic, quasiperiodic, and chaotic behavior; entrainment and synchronization—have found applications in the analysis of many different living systems, from gene regulatory networks to neurons to ecosystems. While ecology was the traditional source of nonlinear dynamics problems, genetic networks have formed a productive modern nexus, connecting the biological physics community to nonlinear dynamics, but also to the systems and synthetic biology communities (Chapters 6 and 7).
While stability and instability are familiar concepts, nonlinear dynamics identified an intermediate class of excitable systems. The prototypical example is the dynamics of neurons: In response to a brief injection of current, the voltage across the membrane of a neuron typically returns to its initial value, but if the current is large enough there will be a large amplitude transient that takes on a nearly stereotyped form—the action potential. Similar behaviors are seen in very different systems, such as the signaling systems that slime molds use to communicate as they make the transition from living as isolated cells to forming an aggregate. Such spatially extended excitable media support novel excitations, such as spiral waves (see Figure 5.6). In some regimes the heart also acts an excitable medium, and spiral waves can be triggers of tachycardia and fibrillation. More generally, models of the heart provide problems for nonlinear dynamics, and this interaction has progressed to the point where it is not unreasonable to think of certain heart conditions as disorders of dynamics.
The classical nonlinear dynamical problems of ecology are low dimensional, from oscillations in predator/prey systems to chaos in seasonal population growth. These problems remain important, not least because these models are actually used to manage fisheries and other human food sources. But in many ways the frontier has shifted to high-dimensional systems, ecologies in which many different kinds of organisms are present simultaneously. This is what is seen in many environments, but there are questions about how this is possible: Why doesn’t the single fastest-growing species win out? There are many ideas about how ecological diversity is maintained: There can be multiple resources, with different species becoming more or less specialized, there can be spatial structures that inhibit competition, or it

could be that persistent diversity is not a steady state but just very slow dynamics. All these ideas, and more, are naturally expressed in dynamical systems language. To make progress often requires some mixing with concepts of statistical mechanics, passing to the limit of very large systems and approximating interactions among species as drawn from ensembles of random matrices. This is a very active field, with theory now being supplemented by large scale, controlled experiments in microbial ecologies.
Finally, what about the connection of biological physics to the physics of the universe as a whole? There are at least two questions. First, in the spirit of cosmol-
ogy, how far back is it possible to trace the history of life on Earth? Second, what would life look like on another planet?
Regarding the deep history of life on Earth, there are classical experiments showing that early atmospheric conditions allowed for the synthesis of moderately complex organic molecules, including amino acids. But there is a huge gap from this to something one might think of as alive. In life today, information passed from one generation to the next is stored in one kind of polymer (DNA), but much of the business of life is carried out by another kind of polymer (proteins). The discovery that some RNA molecules act as catalysts—a function largely filled by proteins today—suggested that there was an early “RNA world.” The first living organisms might have been self-replicating polymers, but there are active efforts to understand how protocells might form, allowing for compartmentalization and encapsulation of an RNA-based genome. While these ideas remain in many ways very speculative, there are serious efforts to map, for example, the landscape of catalytic function versus sequence in RNAs of modest size. This has much in common with ideas about sequence/structure mapping in families of proteins (Chapter 3), and would provide some foundation for more rigorous discussion. In a different direction, several groups are trying to construct artificial cells with limited numbers of components, perhaps providing models for the origin of life but certainly creating a path to understanding by building.
Taking the analogy to cosmology seriously, one might say that we have many ideas about the Big Bang, but what is missing is the cosmic microwave background: What can be measured that carries the fingerprints of those earliest moments? By sequencing the genomes of more and more of the different organisms alive today, it seems possible to construct an evolutionary tree that points to common ancestors further and further back in time. But this assumes that all relationships fit on a tree, and this is not quite true. Bacteria can exchange genetic material without reproducing, and it has been conjectured that this was more prevalent in the distant past. If this is correct, there was a murky ground of information exchange out of which the modern tree of life grew. At present, there is no way to get an image of this ground.
The question of life on other planets of course is given greater urgency by the 1995 discovery that there are other planets orbiting stars similar to the sun. As more and more such planets are discovered, the range of conditions comes closer and closer to those which seem hospitable for life. What should we expect to see? Measurements will be observations from afar, so what is needed are essentially spectroscopic signatures. It is unreasonable to think that life on Earth is so special that all possible life forms will use the same molecules. Recent work suggests focusing instead on the complexity of molecular ensembles: To be alive is not to have one crucial molecule, but rather to have many species of molecules connected by a reaction network. In this direction, it is important to understand whether the
molecules of life on Earth form a network with special properties, distinguished from a random jumble of molecules of similar size and elemental composition.
Perhaps the most important result of thinking more concretely about how to search for life on extra-solar planets is that we are not so sure what features of life on Earth constitute its defining physical characteristics.














