
5
The Life-Course Contribution to Ethnic Disparities in Health
Clyde Hertzman
Do the following observations belong together?
-
Observation 1: Protective early life factors
Birth cohort studies have shown that early life factors contribute to the risk of coronary heart disease, non-insulin-dependent diabetes, obesity, elevated blood pressure, age-related memory loss, and schizophrenia later in life (Barker, 1992, 1994, 1997; Barker and Osmond, 1986; Blane et al., 1996; Davey Smith, Hart, Blane, Gillis, and Hawthorne, 1997; Davey Smith, Hart, Blane, and Hole, 1998; Eriksson et al., 1999; Frankel, Elwood, Sweetnam, Yamell, and Davey Smith, 1996; Kuh and Ben-Shlomo, 1997; Marmot and Wadsworth, 1997; Osmond and Barker, 2000; Ravelli, Der Meulen, Osmond, Barker, and Bleker, 1999; Ravelli, Stein, and Susser, 1976; Susser et al., 1996).
-
Observation 2: Birthweight differences
In a recent study of birthweights among U.S.-born blacks, African-born blacks, and U.S.-born whites in Illinois (1980-1995), the mean birthweight of 44,046 infants of U.S.-born whites was 3,446 grams; of 3,135 infants of African-born blacks, 3,333 grams; but of 43,322 U.S.-born blacks, only 3,089 grams (Davis and Collins, 1997).
-
Observation 3: Health status differences
Another recent study (Fang, Madhavan, and Alderman, 1996) showed that the health status of blacks living in New York City as
-
adults was associated with their place of origin: the northern United States, the southern United States, or the Caribbean. The health status of those from the Caribbean was similar to New York City whites, and better than those from the northern United States; in turn, those from the northern United States had better health status than those from the southern United States.
-
Observation 4: Infant mortality, Aboriginal population
Infant mortality among the Aboriginal population in British Columbia (BC), Canada, is substantially higher than the non-Aboriginal population. At the same time, life expectancy is lower than the rest of the BC population. However, what is most striking is that, in both cases, there are very large regional differences among Aboriginal populations residing in different parts of the province—variations much larger than for the non-Aboriginal population. As shown in Figures 5-1 and 5-2, the regions with the “best” Aboriginal health status are close to, or overlap with, the
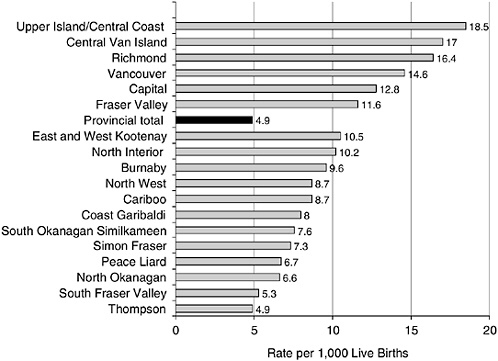
FIGURE 5-1 Infant mortality by health region for status: Indians, British Columbia, 1991-1999.
SOURCE: British Columbia Vital Statistics Agency (2001).
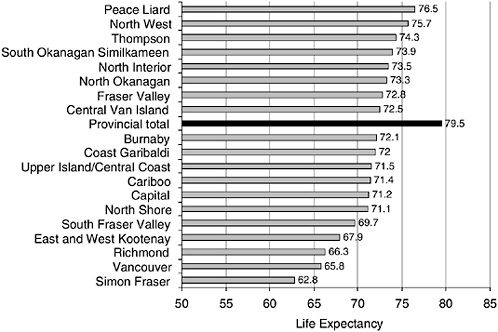
FIGURE 5-2 Life expectancy by health region for status: Indians, British Columbia, 1995-1999.
SOURCE: British Columbia Vital Statistics Agency (2001).
-
overall population average (British Columbia Vital Statistics Agency, 2001).
-
Observation 5: Teenage suicide rates, Aboriginal population
Rates of suicide among Aboriginal teenagers in BC are, on average, much higher than the non-Aboriginal population. However, when the 196 reserve communities across BC are grouped according to the degree of local control they currently exercise over land, health, education, cultural, and other governmental services, the rate of teen suicide falls monotonically as the number of factors under local control goes up (Figure 5-3), such that the teenage suicide rate on reserves that control all these factors is lower than the non-Aboriginal rate for the province (Chandler and Lalonde, 1998).
This chapter contends that these observations, although diverse, form a consistent pattern that is comprehensible from a population health perspective and that can help to explain ethnic disparities in health status.
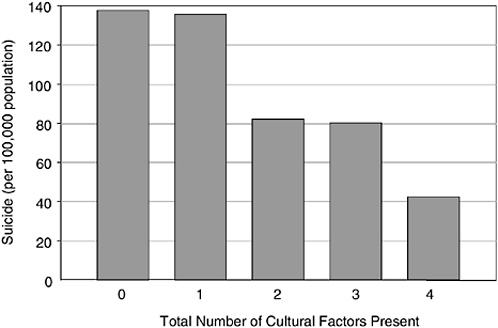
FIGURE 5-3 Aboriginal youth suicide rates by number of factors present in the community.
SOURCE: Chandler and Lalonde (1998).
POPULATION HEALTH PERSPECTIVE
Regarding the life-course dimension of ethnic disparities in health, the population health perspective begins with an incumbent hypothesis: that systematic inequalities in health status among distinct ethnic groups within a society are a product of the interaction between the developmental opportunities and vulnerabilities at each stage of the human life-course, on the one hand, with widely varying attributes of experience at the intimate, civic, and socioeconomic levels of society, on the other. Developmental stages are universal across human populations, notwithstanding ethnicity. In contrast, the nature of day-to-day life experience may differ systematically among ethnic groups. Systematic differences in health status emerge, primarily because some life-course interactions with life experiences are more health enhancing than others. The hierarchy of health status among different ethnic groups in society reflects the rank order of the average quality of life-course/societal interaction experienced by each ethnic group.
According to this hypothesis, ethnic differences in health status emerge from the same sort of nature-nurture interactions that produce socioeconomic differences in health status across the life-course. To produce systematic differences in health status, ethnic differences must be associated with systematically different developmental experiences, and systematically dif-
ferent “qualities” of interaction between the developing individual and his or her intimate, civic, and socioeconomic environment.
From this perspective it is possible to see how the following five introductory observations fit together:
-
Observation 1: Protective early life factors
-
Observation 2: Birthweight differences
-
Observation 3: Health status differences
-
Observation 4: Infant mortality, Aboriginal population
-
Observation 5: Teenage suicide rates, Aboriginal population
The first three observations listed are examples of the protective effects of early experience: those who grew up in a society where they were not a stigmatized ethnic minority enjoyed health protection, both in adulthood and in the intergenerational transmission of birthweight. Those with the same racial/ethnic background, who grew up as a stigmatized minority in a white-dominated society, were not similarly protected. These examples, of course, are potentially confounded by differential migration according to health status, differential access to health care, and residual genetic/constitutional differences within nominally common “racial” groups (to be discussed).
The latter two observations listed are examples which deal exclusively with nonmigrants (indeed, “original peoples”) in a jurisdiction with no financial barriers of access to medical care. Moreover, the Aboriginal peoples of BC fall into several different “nations” that, in turn, have different customs and traditions and, because of the physical barriers of mountains and ocean, had little tendency to interbreed before the era of modern transportation. Despite this, variations in health status across the province do not follow any known intra-Aboriginal national pattern, and huge differences by region are the rule. The nearly fourfold variation in infant mortality shown in Figure 5-1 is also unrelated to proximity to medical care. This range of mortality is a proxy for variations in the “quality” of the reproductive and early child environments encountered by the Aboriginal population. The teenage suicide example is the most direct for our current purposes. The degree of local control of the civic environment in Aboriginal communities is a measure of their liberation from external domination by white society. Figure 5-3 shows that the “degree of liberation” has a dose-response relationship to a key outcome of “healthy child development”: teenage suicide.
UNDERSTANDING THE LIFE-COURSE
Life-course factors are now well understood to affect a diverse range of outcomes, from general well-being to physical functioning and chronic
diseases (Keating and Hertzman, 1999; Kuh and Ben-Shlomo, 1997; Marmot and Wadsworth, 1997). Exposure to both beneficial and adverse circumstances over the life-course will vary for each individual and will constitute a unique “life exposure trajectory” that will manifest itself in different expressions of health and well-being.
Although this general perspective is noncontroversial, there are large problems of operationalization. Perhaps the most significant is the way in which early life phenomena are studied under the rubric of “development” whereas later life phenomena are studied under the rubric of “aging.” Unlike the shorter lived primates, whose complete life-course and those of their offspring can be studied during a “normal” 30-year research career, human longevity poses a problem of how to “stitch together” longitudinal observations from early, middle, and late life into a coherent and credible whole. Because this process of stitching together has not been taken very far, development and aging research often appear to be in conflict. The output of developmental studies of health is often interpreted to claim that, by a certain (undefined) age, future health status has been predetermined, and the rest of the life-course merely reveals what the early years have fated. On the other hand, aging research often seems to treat the individuals who enter middle-aged and elderly cohorts as blank canvases on which later life events will be imprinted.
These separate solitudes need to be brought together. The next several paragraphs present the determinants of health from a developmental perspective, and will appear to some readers as though they reinforce the schism just described. To compensate for this weakness, the succeeding paragraphs will try to connect the developmental perspective to the aging perspective.
Biological Basis
The biological basis for life-course relationships to health arises from the similarity of human development to that of other primate and nonprimate species. From these sources we know that brain structures and functions can develop differently in differing early environments. The developing brain is an environmental organ that sculpts itself according to experience (Cynader and Frost, 1999). Differential experiences are closely associated with systematic differences in the function of at least three important physiological control systems: the HPA axis (the hypothalamic-pituitary-adrenal system, which controls cortisol secretion), the SAM axis (the sympatho-adrenal-medullary system, which controls epinephrine and norepinephrine secretion), and the PNI axis (the psycho-neuro-immune axis; that is, central nervous system influences on immune system function and blood-clotting factors). The SAM, HPA, and PNI axes all have a “life” within individuals in society, which, in turn, has an empirically demonstrable role in producing systematic differences in health over the life-course
(Hertzman, 2000). The hypothesis here is that early life experience profoundly shapes the ways these axes develop; that these body-regulatory and defense systems, in turn, experience the environment and respond to it in radically different ways in different individuals according to the differences in their early experiences; that these responses influence the lifelong health of the individual; and finally, that systematic differences in these responses across population groups in a society (whether defined by socioeconomic status, ethnicity, or geography) contribute to health disparities. The interest here is in the interplay—day to day, hour to hour, moment to moment—between the environments where people live, work, and grow up, on the one hand, and the development and responses of the SAM, HPA, and PNI axes, on the other, which lead to systematically differing health expectancies over a lifetime.
There are, of course, an infinite number of ways in which exposure to expression relationships can unfold over time. In some instances, there will be contemporaneous exposure and expression. If a piano falls on one’s head, the exposure and the effects on health and well-being are simultaneous, unequivocal, and easily measured. But in most cases the connections from exposure to expression will be more subtle and will play out over long stretches of the life-course. The possible long-term exposure to expression relationships cluster into three generic patterns: “latency,” “cumulative,” and “pathway” (Hertzman and Wiens, 1996; Hertzman, Power, Matthews, and Manor, 2001; Power and Hertzman, 1997).
Latency
“Latency” means a relationship between an “exposure” at one point in the life-course and the probability of health “expressions” years or decades later, irrespective of intervening experience. The following examples validate the concept of latency.
Barker’s “fetal origins hypothesis” posits that several chronic diseases in adult life originate from permanent changes in the structure, physiology, and metabolism of the fetal body as an adaptation to undernutrition (Barker, 1997). Studies of British records from early in the 20th century show associations of birthweight, placenta size, and weight gain in the first year of life with cardiovascular disease in the fifth decade (Barker, 1992, 1994). Fetal growth also has been associated with the development of insulin resistance in mid-life and onset of diabetes in the sixth and seventh decades of life (Osmond and Barker, 2000). Evidence of the relationship between birthweight and cardiovascular risk factors, such as hypertension, has now been found in numerous populations (Huxley, Shiell, and Law, 2000).
Studies of the Dutch Hunger Winter (1944-1945) showed that maternal nutritional deficiency in pregnancy was associated with an increased risk of
antisocial personality disorder (ASPD) and schizophrenia among offspring in adult life (Neugebauer, Hoek, and Susser, 1999; Susser et al., 1996). These effects appeared to depend on the timing of prenatal insult: men exposed to severe maternal nutritional deficiency during the first and/or second trimesters of pregnancy had a risk of ASPD that was 2.5 times that of men who had not been exposed prenatally (Neugebauer et al., 1999).
Perhaps the quintessential example of latency arises from longitudinal studies, not of humans, but of rats. In several rat species, the degree of early suckling and licking of the newborn by its mother has a lifelong effect on a range of functions, most notably stress response, learning, and memory (Francis, Diorio, Liu, and Meaney, 1999). Maternal licking and suckling early in life influence the expression of certain genes that regulate the development of the HPA axis, which regulates the amount and pattern of cortisol that the rat’s body produces in response to stressful life circumstances. Cortisol has a wide range of effects on physiological functioning. In the short term, it makes the rat more alert and ready to cope with danger. Over the long term, however, high levels of cortisol have the effect of accelerating the aging process in various organs within the body (Sapolsky, 1992). Infrequently suckled baby rats tend to develop, permanently, a highly reactive HPA axis. Frequently suckled rats develop a “down-regulated” HPA axis. The implications of these differences are far reaching. Having a down-regulated HPA axis is associated with better decision-making functions, quicker task learning, and a slower rate of loss of learning and memory functions. Thus, the frequently suckled baby rats acquire a lifelong advantage as a result of a systematic difference in early nurturant experience (Francis et al., 1999).
Cumulative Effects
“Cumulative” refers to multiple exposures over the life-course that have combined effects on health. Vulnerability and/or resiliency can combine over time through cumulating injuries/risk factors or cumulating privileges/protective factors. In some cases, cumulation will occur in a dose-response manner. For example, duration of exposure to adverse socioeconomic circumstances affects childhood growth, with persistent poverty increasing the risk of childhood stunting (low height for age) and wasting (low weight for age), whereas single-year income measures do not (Miller and Korenman, 1994).
Duration of exposure to particular socioeconomic circumstances shows a cumulative effect on several health outcomes in adulthood. In the 1958 British birth cohort, the occupational class of the male parent over the first three decades of life was found to be strongly predictive of health status in early adulthood (Power, Manor, and Matthews, 1999). Figure 5-4 shows a dramatic direct relationship between the risk of poor health and a score
that is constructed by combining occupational status at four points in the life-course (with the lowest score indicating continuous high occupational status, and the highest score the reverse). The lifetime score represents cumulative duration and intensity of material/social privilege or deprivation. Those who were always in the lowest occupational class were approximately four times as likely to report poor health as those who were always in the highest. “Lifetime” occupational class is a stronger predictor of poor health than occupational class at any single point in time, indicating that duration of exposure to poor socioeconomic circumstances matters a great deal. Moreover, for both men and women, the risk increases monotonically with increasing lifetime scores (Figure 5-4). In other words, circumstances at each life stage build on one another. Similar patterns have been found by others (Lynch, Kaplan, and Shema, 1997).
Another variant would see different factors over the life-course add to effects of factors from early life. This might be described as a “multiple factor additive” variant. In the 1958 British birth cohort, chronic illness and disability at age 33 were predicted by factors in early life (childhood socioeconomic disadvantage and height), in adolescence (behavioral adjustment), and in adult life (injury and underweight/overweight) (Power, Li, and Manor, 2000).
Pathways
Finally, “pathways” refer to the ways in which individuals get onto well-worn life-course trajectories that carry with them a set of “health
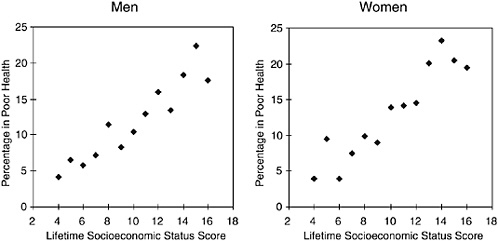
FIGURE 5-4 Poor health includes subjects who rated their health as fair at age 33 and cumulative socioeconomic circumstances (birth to age 33).
SOURCE: Power, Manor, and Matthews (1999).
chances” based on the socioeconomic and psychosocial circumstances that prevail along them. This occurs because a sequence of experiences at one stage of the life-course will influence the probability of other experiences later in the life-course. For those using different terminology, it should be recognized that social mobility is a function of the pathways model. There is a dominant sequence of events that unfolds as follows:
-
At first, early life social origins influence readiness for school (Ross and Roberts, 1999). Studies among preschool-age children in Canada and the United States have demonstrated that the circumstances of the family early in the child’s life (income, education, single parenthood, and parenting style), neighborhood (safety, cohesion, and socioeconomic ghettoization), and social institutions (access to “quality” child care arrangements) are powerful determinants of cognitive and behavioral development (Duncan and Brooks-Gunn, 1997, 2000; Duncan, Brooks-Gunn, and Klebanov, 1994; Kohen, Hertzman, and Brooks-Gunn, 1998).
-
Cognitive, emotional, and behavioral readiness for school have consequences for school success and social adjustment. School readiness, as measured by cognitive and social-emotional competencies, is important because children who are not ready for school are more likely to experience school failure, conditions of unemployment, criminality (Tremblay, 1999; Tremblay, Masse, Perron, and LeBlanc, 1992), and psychological morbidity in young adulthood (Power et al., 1991). Intervention studies have demonstrated that improvements in the childhood environment can provide a long-lasting benefit to an individual’s life-course trajectory (Hertzman and Wiens, 1996). The Perry Preschool Project demonstrated that an intensive child care and home visiting intervention at ages 3 and 4 could improve high school graduation rates, reduce teenage pregnancy, increase income, reduce dependence on social services, and reduce criminal activity by age 27 among socially disadvantaged children. From the standpoint of the pathway model, the most interesting observation was that the long-term benefits seemed to accrue from the manner in which the intervention improved the children’s transition to school (Schweinhart, Barnes, and Weikart, 1993).
-
Next along the pathway, there is abundant evidence that people with a poorer education have more harmful health-related behaviors than those with a better education. Evidence for this pattern is found in relation to smoking, diet, wearing seat belts, and seeking preventive health care, such as immunization and PAP smears (Davey Smith et al., 1998; Winkleby, 1992).
-
Adult socioeconomic circumstances, also closely associated with educational attainment, have an impact on health in adult life (Drever and Whitehead, 1997; Evans, Barer, and Marmor, 1994; Kunst, Geurts, and van
-
den Berg, 1995; Kunst, Groenhof, Mackenbach, and Health, 1998). International studies of European, American, and Canadian populations demonstrate poorer self-reported health and mortality of the least educated groups (Feldman, Makuc, Kleinman, and Cornoni-Huntley, 1989; Kunst et al., 1995; Pappas, Queen, Hadden, and Fisher, 1993). Thus, social origins can have their effect on health status through their influence on social destinations.
Figure 5-5 (adapted from Hertzman et al., 2001) is meant to illustrate how latent, cumulative, and pathway effects combine with current circumstances to help “explain” health status across the life-course. The arrow in the figure represents the life-course, intersecting with the bulls-eye that represents society. Society is illustrated by three concentric circles representing increasingly broader levels of social aggregation where determinants of health are found: the personal social support network (most intimate), civil society (intermediate), and the socioeconomic environment (broadest). Despite problems of visual representation, the arrow is meant to intersect the bulls-eye at the most intimate level, and does so continuously on a day-to-day basis throughout the life-course. Thus, the figure represents the idea that health status is an emergent property of the ongoing interactions between the individual (at each stage of development across the life-course) and the conditions they encounter in the intimate, civic, and broader socioeconomic environments.
The example illustrated in Figure 5-5 uses this model, in a somewhat reduced form, to display findings from the 1958 British Birth Cohort study. In particular, it shows how experiences during the first 16 years of life
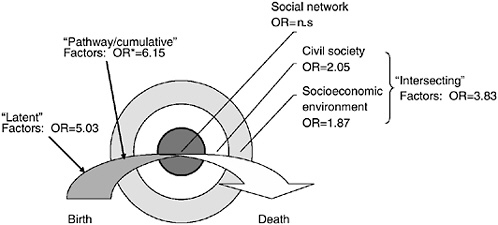
FIGURE 5-5 Contributions to self-rated health at age 33, 1958 birth cohort. OR = odds ratio. For interpretation, please see text. n.s. = not significant (p > 0.05).
affect self-rated health by age 33. In this model, the latent effects were the proportion of adult height the child had obtained by age 7, the regularity with which parents read to the child up to age 7, and the degree of socioemotional adjustment to school by age 7. These variables represent the three principal dimensions of early child development: physical, cognitive, and social-emotional-behavioral. In the model they were statistically independent of one another and of factors that came later in the life-course. The odds ratio of 5.03 suggests that those who have relatively rapid early growth, who were read to consistently, and who adjusted easily into school were one-fifth as likely to be in a state of fair or poor health by age 33 as those who grew slowly, were not read to consistently, and had trouble adjusting to school.
The statistically independent cumulative and pathway effects were: cumulative socioeconomic status at age 0, 7, 11, and 16; socioemotional adjustment to school at ages 11 and 16; and end-of-school qualifications. The model suggests that those who were always in the lowest socioeconomic category throughout childhood, who had ongoing trouble adjusting to school, and who dropped out without formal qualifications were approximately six times more likely to be in a state of fair or poor health by age 33 than those with the opposite status on each of these factors.
In addition, the model shows the extent to which current circumstances, at age 33, contribute to health status above and beyond the contributions of the earlier life-course. The odds ratio for fair/poor self-rated health is 1.87 comparing the lowest and highest current social class; 2.05 for those least affiliated to most affiliated in their local communities; and 3.83 for those with the least sense of control at work and in life, compared with those with the most.
I am unaware of analyses that parallel these by ethnic group in any of the world’s large birth cohort studies. In particular, none of the American birth cohort or child longitudinal studies have attempted this sort of analysis. A great deal of work has been done trying to explain educational and developmental differences by nominal ethnic category in the United States, but little of this work has been further linked to health status in adulthood. I would propose that this be a priority to help understand the life-course contribution to ethnic differences in health status in the United States.
BRINGING DEVELOPMENT AND AGING TOGETHER
So far, this analysis covers only the period from birth to early adulthood. How does the developmental perspective link to aging? Here, I propose that four key points of tangency call out for further empirical investigation.
The Shakespearean Connection
In the soliloquy “All the World’s a Stage,” Shakespeare presented to the Western world a story of the human life-course as a rise (during early life) and a fall (during late life). Although there are more humanistic ways to view the life-course (such as the traditional Japanese notion of life being a voyage that ends at a final destination), the Shakespearean model is more suited to the biological realities. In this model, development is the rise and aging is the fall. Thus, the health and well-being of a given individual on the downward slope of the life-course would, in principle, be determined by two parameters: first, the height at the top of the rise—that is, how successful the process of development had been; and second, the rate of decline along the downward slope of aging. From this perspective, then, development and aging are tied together by the accumulation of “reserve capacity” during development and also by the prospect that early developmental experiences may influence the later life rate of decline.
Similarity of Life-Course Processes
Latent, pathway, and cumulative processes apply late in life, just as they do in early life. The example of the rate of cognitive decline in late life, the incidence of dementia, and their association with level of education earlier in life is a case where all three processes are relevant (Evans et al., 1993). Those with greater levels of education early in life may have slower rates of cognitive decline later in life because of a neurochemical reserve brought about as a result of the education process (a latent effect); or due to the social mobility/enhanced self-esteem associated with greater education (a pathway effect); or due to an association between greater education and greater cognitive stimulation throughout the working life (a cumulative effect).
Similarity of the Determinants of Healthy Aging and Healthy Child Development
Socioeconomic status, social integration, social support (Seeman and Crimmins, 2001), self-esteem (Seeman et al., 1995), and a sense of control or powerlessness at work and in life (Seeman and Lewis, 1995) have all been shown to influence the aging process, either through direct effects or through buffering or exacerbating of existing vulnerabilities. Many of these factors are also important influences in early life, and are reflected in Figure 5-5. Thus, notwithstanding the “rise and fall” dichotomy, the factors that affect the height of the rise and the rate of the fall may well be held in common.
The Life of the Host Defense Systems in Human Society
In our work on the early life-course, we have coined the term “biological embedding” to refer to the process whereby systematic differences in life experience translate into systematic differences in the response of the body’s host defense systems to stress that, in turn, will affect organ-system aging and health status in later life (Hertzman, 2000). In the aging field, a highly complementary concept of “allostatic load” has been developed and operationalized to a much greater extent than biological embedding. Allostatic load is composed of a group of metabolic variables that, taken together, represent the metabolic set point of each aging individual’s defense/stress response system (Seeman and McEwan, 1996). Allostatic load, stress, and aging can then be likened to a spring. As a spring is repeatedly used, its position at rest may become altered, altering its response to force bearing, and reducing its elasticity over time. In humans the metabolic set point (allostatic load) interacts with environmental stressors (analogous to force bearing). If allostatic load is not optimal, physiologic elasticity (that is, the ability to mount an effective physiological response to stress and return to baseline) may be lost at an accelerated rate, leading to accelerated biological aging. The key point of tangency, and the key issue for life-course research, is the extent to which the process of biological embedding can “explain” individual or group differences in allostatic load, as they are first measured in adulthood. The leading hypothesis would be that adverse early experiences would lead, through biological embedding, to a high-risk allostatic load in mid-life and late life.
OPPORTUNITIES FOR ETHNICITY TO AFFECT DEVELOPMENTAL TRAJECTORIES
Latency and Ethnicity
In theory, there are three ways that ethnic differences could translate into latent effects on health across the life-course. First, there is the potential for prenatal transmission of socioeconomic or psychosocial stresses through transplacental transmission of maternal experience via the secretion of cortisol and other stress hormones. The hypothesis here is that pregnant women from relatively deprived ethnic backgrounds will be more likely to find themselves under some form of uncontrollable socioeconomic or psychosocial stress during pregnancy. It has been shown that increased HPA axis activity during pregnancy can be reflected in increasing circulating cortisol in the fetus, both in humans and nonhuman primates (Gunnar and Barr, 1998; Post and Weiss, 1997). Fetal cortisol, in turn, has been shown to be active in neurobiological development. In Coe’s (1999) work on rhesus
monkeys, a randomized, controlled trial of increasing the level of uncontrollable maternal stress for short bursts during pregnancy (keeping all other aspects of gestational environment constant) led to developmental delays in their offspring. The Dutch Hunger Winter studies and several studies of the risks of low-birthweight and small-for-gestational-age babies have demonstrated that prenatal stress can affect birth outcome. However, I am unaware of any studies that directly connect ethnicity, circumstances during pregnancy, transplacental transmission of maternal experience, and developmental outcome.
Second, there is the potential for ethnic differences in parenting style to make a difference in early cognitive and social development. It is well known, and reasonably well documented, that different ethnic groups tend to have different bonding and attachment styles with their newborns; different values and beliefs regarding when and how to talk with young children and read to them; and parenting styles that can vary from the interactive to the apathetic to the authoritarian. It is now clear that each of these differences can make a difference in the cognitive and social development of the child. For example, it has been shown that the acquisition of vocabulary follows a dose–response relationship according to the degree of oral fluency in the home (Hart and Risley, 1995). I am not aware of any systematic measurements of ethnic differences in oral fluency and reading in the home. Anecdotally, however, these are thought to be large in Canada, with Aboriginal homes believed to be relatively lacking in verbal interchange.
In the Canadian National Longitudinal Survey of Children and Youth, apathetic and authoritarian parenting styles were strongly associated with developmental delay (both social and cognitive), even after other factors were taken into account (Willms, 2000). Once again, we have little useful information on how parenting styles vary by ethnicity. At the forefront of research in early language acquisition is the potential synergy between language and social development, and the degree to which visual contact with a parent or caregiver during language acquisition helps define its social meaning. This latter may be very relevant to ethnicity and early development. For example, Bornstein and Cote (2001) have shown that different cultural groups around the world tend to orient their babies differently when they are being held and carried. In North America, we tend to orient babies outward toward the world, whereas in Japan, they tend to face inward toward the parent.
Finally, there is the prospect that ethnicity is associated with transmission of systematically different messages regarding one’s place in the social hierarchy during the neurobiological developmental phases of host defense and higher executive functions. The primitive structures in the mid-brain that sense and initiate physiological responses to threat develop over the first few years of life and, between the third and tenth year of life, form
extensive synaptic connections with the executive functions of the prefrontal cortex. Both the early conditioning of the mid-brain and the nature of the connections to the prefrontal cortex appear to be highly conditioned by the environment in which the child develops. At present, we do not know the extent to which parents’ perceptions of being a stigmatized group within society can be transmitted to the child during early development or the extent to which a strong family network can buffer such putative effects. We do know that children who grow up in physically abusive environments tend to have a lowered threshold for the recognition of, and physiological reaction to, threat in adult faces (Pollak, Cicchetti, and Klorman, 1998). This is evidence, albeit fragmentary, that early transmission of place in the social hierarchy can occur and can endure.
Children growing up in unsafe, noncohesive, and low socioeconomic ghetto neighborhoods tend to reflect these conditions in cognitive and social development delays by age 4 (Kohen et al., 2002). In Canada, Aboriginal children tend to grow up in such neighborhoods and in the United States black and Latino children tend to do so as well. But once again, the principal research question is unanswered: To what extent is ethnicity associated with differences in early childhood environments that condition key sensing and executive response systems in ways that influence health and well-being across the life-course? One excellent research opportunity here would be a life-course study of the factors that contribute to higher blood pressure among American blacks than whites.
Pathways and Ethnicity
The story of pathways and ethnicity is, first, the story of what happens when a visible ethnic status becomes a social constraint (or social opportunity). The ways in which visible status can affect educational and social mobility through the perceptions and actions of powerful individuals and institutions are too numerous and well known to review here. Insofar as these perceptions have been prejudicial to visible minorities, limiting their trajectories of income, education, and/or occupation, they can be taken to be relevant determinants of ethnic differences in health status, because adult socioeconomic status carries powerful health expectations with it.
The second large factor is readiness for school. When systematic differences in early childhood experiences lead to differences in children’s readiness for school, a negative pathway effect is probable. It is well known that children who enter school developmentally unready are themselves at risk of school failure. Classroom composition also appears to affect the educational trajectories of those who come into school fully ready for the experience. Our work in Vancouver shows that the proportion of kindergarten children who are developmentally vulnerable entering school varies sixfold
among neighborhoods. Our data suggest that a high proportion of children not “ready for school” seem to hold back the “ready-for-school” children (Mustard and Picherack, 2002). Presumably, this is a classroom pacing effect. In Vancouver, the neighborhoods most heavily populated by Aboriginal children are those with high rates of crime, commercial sex, and drugs; high proportions of children entering school in a developmentally vulnerable state; and low levels of subsequent educational success. Although similar data on readiness for school are yet to be collected in American cities, it is well known that the neighborhoods heavily populated by Latino and black children have similar characteristics to those populated by Vancouver Aboriginals.
Other aspects of the pathway effect include the role of parental expectations on children’s developmental trajectories and local levels of environmental enrichment: neighborhood programs supporting parenting and development, school quality beyond classroom composition, and the availability of successful role models during the teenage years and adulthood. Sweden and Japan have been successful in compressing socioeconomic gradients in achievement of basic literacy and numeracy skills by introducing flexibility into the transition from preschool to school. In Sweden this is accomplished through a flexible transition from play-based to formal learning styles. In Japan it is based on flexible teaching and learning styles in the primary grades, especially with respect to quantitative concepts.
It should be reemphasized here that early life conditions affect social mobility through cognitive, social-emotional-behavioral, and physical readiness for school according to the pathway model. To the extent that ethnic differences in school readiness exist, and are related to early life experiences, there will be a social mobility effect (or, for that matter, a social immobility effect) on later health status. To the extent that ethnicity differentially affects early physical development, subsequently affecting school performance and social mobility, there could be a reverse causality effect.
Cumulative Effects and Ethnicity
The most obvious cumulative effect is that associated with the duration and intensity of socioeconomic deprivation among those ethnic groups who tend to be deprived. Here, the literature on socioeconomic deprivation and health per se can simply be read into the record. However, a variety of other “positive” cumulative effects may apply to specific ethnic groups. In Texas first-generation Mexican families have much better health status than would be expected based on their socioeconomic conditions. This has been attributed, in passing, to the protective effects of belonging to a strong, traditional extended family that, after a generation in America, begins to break
down. There is also a well-known phenomenon in Canada and the United States of increased late-life survivorship among those ethnic groups whose early life survival rates are relatively low, particularly a tendency for supplementary life expectancy beyond age 70 to be higher among African Americans and Canadian Aboriginals than the white population, despite their lower life expectancy at birth.
A second prospect, which undoubtedly will be receiving increased attention in coming years, is disjunction between genetic susceptibilities and environmental exposures. This can become relevant either when an ethnic group migrates to a new place with a mixture of biological, physical, chemical, cultural, and nutritional characteristics to which it has not been historically adapted or when a host population is overtaken by an in-migrating population that gains hegemony with respect to the biological, cultural, nutritional, or chemical environment. The roles of smallpox and tuberculosis in wiping out host populations in North America during the early years of European colonization are examples of the latter. The spread of syphilis to the European population and the loss of the adaptive value of sickle cell trait to malaria among the American black population are examples of the former. As of this writing, a key issue in Canada is the role of Western diet in the epidemic of diabetes mellitus among the Aboriginal population. Traditional Aboriginal diets were largely absent of unrefined carbohydrates, so the leading hypothesis is that the increasing penetration of Western diet has created an invidious interaction between diet and genetically determined metabolic processes in Aboriginals that are easily overwhelmed by large carbohydrate loads. However, it is likely that, like most chronic diseases, the full explanation will turn out to be more multifactorial in nature.
COMPLEMENTARY EXPLANATIONS FOR ETHNIC DIFFERENCES ACROSS THE LIFE-COURSE
Four groups of explanations could potentially account for ethnic differences in health status across the life-course through population selection processes that are not usually considered to be life-course phenomena. I label them differential migration, differential social mobility, genetic mono-causation, and differential utilization of health services. This section will briefly show how difficult it is, in practice, to make a clear-cut distinction between life-course and non-life-course phenomena. Something that at first glance appears to “confound” life-course may actually be a contributor to it.
Differential Migration
Take, for example, the differential migration hypothesis: Populations that migrate around the world seeking freedom and opportunity are sys-
tematically different from both the populations they left behind and those that they encounter when they arrive in their new homes. Also, populations that move voluntarily are systematically different from those that are subject to forced migration. They, in turn, are systematically different from those they leave and those they encounter when they arrive at their involuntary destination. A wealth of evidence supports the existence of each of these phenomena from around the world. Moreover, the first two observations, protective early life factors and birthweight differences, given in this chapter could be accounted for by the fact that Caribbean and African blacks in the United States are voluntary migrants, whereas American-born blacks are either nonmigrants or, if ones’ historical memory is long enough, forced migrants.
As a “non-life-course” hypothesis, differential migration has the potential to conceal important information by avoiding the question of how migrants differ from the rest of the population and how these differences evolve over time. For example, there is evidence that over the generations, the children of black Caribbean immigrants to the United States start to develop an American black “racialized” identity that strips them of certain beneficial effects of their parents’ identities (Waters, 1999). In other words, what appears at first to be a characteristic of the migrant may be a characteristic of the migrant-environment interaction during vulnerable phases of social development over the life-course. Another example of differential migration revealing a life course dimension concerns cancer in Japanese migrants to California. They arrived in California with the typical Japanese ratio of stomach-to-colon cancer risk that, within two generations, converted to the typical North American pattern (Doll and Peto, 1981). By following migrant families over several generations, it was possible to conclude that an environmental/life-course interaction was decisive in what, initially, looked like a fixed population difference between migrant and host population.
Differential Social Mobility
Differential social mobility is well known to be influenced by early social and economic factors shaping the life-course trajectory. However, critical appraisal requires consideration of the prospect that differential mobility may also be based, to some extent, on biological characteristics differentially distributed by ethnicity that make some groups more (or less) adaptable to the socioeconomic and psychosocial environments of modern society. This hypothesis would suggest that the social destinations of different ethnic groups are (to some extent) predetermined, and thus the health characteristics of these social destinations are predetermined, too. This hypothesis rests on old and highly politically charged premises that, when articulated, rarely lead to useful investigation.
Whatever the role of predestination, two counterfactuals must be stated. First, the more we know about brain development, the clearer it becomes that the developing brain is an environmental organ and is profoundly adaptable according to early environmental experiences. That is, phenomena that might be seen as static characteristics of a given ethnicity may turn out to be products of interactions between environment and life course. Second, with the passage of decades and centuries, privileged and deprived ethnic groups tend to change places with one another, and with these changes come parallel changes in the image of their capabilities as a group. One relevant example is the radical transformation of the image of Asian peoples in North America during the 20th century.
Genetic Mono-Causation
Genetic mono-causation refers to the effects of genetic differences between ethnic groups that are uninfluenced or modified by the physical or social environment, past or present. Where these exist, they could, in theory, lead to systematic differences in health status among ethnic groups that would be unchangeable over place and time. Although there are plenty of well-known examples of health conditions that are found in disproportionate numbers in particular ethnic groups, these conditions are all comparatively rare and cannot explain large differences in health status by ethnicity. For genetic mono-causation to play a significant role, there would need to be something on the order of a senescence gene (or gene complex) that accelerated or slowed the aging of cells across a variety of organ systems, and it would need to be nonrandomly distributed by ethnicity. Although such genes may exist, and may be distributed in a nonrandom fashion among different ethnic groups, it is worth noting that during the 20th century the average life expectancy of the citizens of the world’s 25 wealthiest countries rose by 30 years, irrespective of ethnic differences and without recourse to genetic change. There is no evidence that genetic mono-causation has played a significant role in explaining the differences in health status among different ethnic groups that exist today, and it is likely that the role of genetic mono-causation will remain marginal in the future.
Differential Utilization of Health Services
Finally, there is the issue of differential utilization of health care services, by ethnic groups, that are unrelated to differences in health status. These certainly exist. Language barriers of access to care have been documented in many jurisdictions and are strongly associated with ethnicity. For example, in the early 1990s in inner-city Vancouver, 25 percent of surveyed parents who had not taken their primary school-aged children to a dentist
in the past year cited “language barriers” as the reason (Hertzman, McLean, Kohen, Dunn, and Evans, 2002). Similarly, cultural factors may inhibit access to care. In Canada, an important subject of investigation and service innovation is ensuring timely access to health care for immigrant women from cultures where women have traditionally played a subordinate social role. Once again, this cannot be construed as a clear-cut non-life-course process because differential utilization of health care early in life can influence health status throughout the balance of the life-course.
AN AGENDA FOR RESEARCH
The framework of understanding presented here strongly implies an agenda for research, and several examples of it have been scattered throughout this chapter. In broad terms, this research agenda hinges on four strategies:
-
There needs to be better exploitation of migrator studies in ways that can estimate the influences of early life factors, social construction of identity, sense of coherence, health selection, and the other categories mentioned. Canada and the United States are two obvious places where such research is feasible because of high rates of immigration, diversity of sources of immigrants, and easy access to second-, third-, and fourth-generation migrant populations. Cooperation must be developed between the research community and the immigration authorities in order to find ethical ways to gain access to sensitive administrative data and direct access to immigrants. In Canada, these relationships are gradually being developed through persistent efforts by population and health researchers to earn the trust of immigration officials for research in the public interest.
-
There needs to be better strategic exploitation of birth cohort studies to understand nature-nurture differences by ethnic status across the life-course. In the United States the primary source for this work is the National Longitudinal Study of Youth (NLSY). It has been used by several investigators cited in this chapter (Duncan and Brooks-Gunn, 1997; Guo and Harris, 2000) to study socioeconomic and neighborhood effects on child development. This data source, in particular, has been used to show how contextual factors have affected differential development among American black and white children. This work needs to be followed forward in time to the ages at which health outcomes become prevalent. To assist this process, it would be of great value to create a strategic partnership with the Panel Study of Income Dynamics (PSID). The PSID is an ongoing annual survey of several thousand households that began in 1968 and has been refreshed over time (Lavis, 1997; McDonough, Duncan, Williams, and House, 1997). It has been a rich source of information on labor market
-
dynamics in relation to adult health outcomes because of its focus on a middle age range. Analysis of early childhood experiences of the PSID cohorts should be carried out in relation to their health outcomes in midlife. This would form a powerful complement to the NLSY and inform it about the direction that adult follow-ups should take.
Moreover, there is a need for American, British, Canadian, New Zealand, and continental European birth cohort studies (and other child longitudinal studies) to develop common protocols for defining different ethnic groups and comparing their life-course trajectories in different societies. This, of course, is easier said than done. Despite the large size of many of these studies, the number of children of a specific ethnic group in any sample may be too small for analysis. Also, developing common international protocols of coordinated analysis runs up against funding asymmetries, researcher resistance, and other factors. UNICEF may have a role here. It has shown a strong interest in child development in different societies, especially those in transition, and may be a powerful platform for brokering useful international, intercohort comparisons.
-
An area in which “quick wins” may be possible would be to construct case studies of successes versus failures of melting pots—working backward from health status to understand why some melting pots support good health status among specific ethnic groups, while others do not. For example, although Canada’s black population is far smaller, as a proportion of the total, than America’s, there has never been a discourse of black health disadvantage in Canada. If Canada-U.S. cooperation like that developed for the recent income inequality and mortality studies were established, it would be possible to subject this to rigorous empirical test, working backward to try to understand any systematic differences found. A similar attempt could be made with Canadian and American Aboriginal populations. Here, we know that although America’s treatment of its native population was harsher than Canada’s in the 17th to 19th centuries, Canada’s policies of forced assimilation were more draconian during the 20th century. Do these differences make a difference with respect to current health status? As it currently stands, it is comparatively easy to capture Aboriginal mortality in Canada, but much more difficult in the United States. Overcoming the obstacles to Canada-U.S. comparisons in this respect should be a priority.
-
Finally, we need to mobilize those who work on the neurobiology, endocrinology, and immunology of human development to understand better how ethnic differences and different forms of ethnicity-society interaction can embed themselves in human biology so that systematic differences in health status can occur. For example, some forms of ethnic-society interactions (material and psychosocial) appear to be harmful to health, and
-
others do not. In another example, it seems to be worse to be black than brown in America despite the visibility of both. Also, it seems that the effects of ethnicity are more pronounced for black men than women in America, at least in terms of economic success and survival through the late teenage years and young adulthood. The field of resiliency (Grotberg, 1995; Werner, 1989) needs to be brought into the discussion here. There is evidence from many manmade and natural disaster zones around the world that a relatively large proportion (approximately 30 percent) of children seem to be strengthened, rather than weakened, by their negative early experiences. They are deemed to be the resilient children. Resiliency seems to accrue to those who have a strong sense of self-efficacy and the sense that they are loved and supported by significant others. Whether or not resiliency can be taught, or engendered through social initiative, may be of great significance in relation to ethnic disparities in health over the life-course and to the biological embedding of adverse early experiences.
REFERENCES
Barker, D.J.P. (1992). Fetal and infant origins of adult disease. London: BMJ Books.
Barker, D.J.P. (1994). Mothers, babies and disease in later life. London: BMJ Books.
Barker, D.J.P. (1997). Fetal nutrition and cardiovascular disease in later life. British Medical Bulletin, 53, 96-108.
Barker, D.J.P., and Osmond, C. (1986). Infant mortality, childhood nutrition, and ischemic heart disease in England and Wales. The Lancet, 1, 1077-1081.
Blane, D., Hart, C.L., Davey Smith, G., Gillis, C.R., Hole, D.J., and Hawthorne, V.M. (1996). Association of cardiovascular disease risk factors with socioeconomic position during childhood and during adulthood. British Medical Journal, 313, 1434-1438.
Bornstein, M.H., and Cote, L.R. (2001). Mother-infant interaction and acculturation: I. Behavioural comparisons in Japanese American and South American families. International Journal of Behavioural Development, 25, 549-563.
British Columbia Vital Statistics Agency. (2001). Regional analysis of health statistics for status Indians in British Columbia 1991-1999. Victoria, Canada: Ministry of Health Services.
Chandler, M.J., and Lalonde, C. (1998). Cultural continuity as a hedge against suicide in Canada’s first nations. Transcultural Psychiatry, 35, 191-219.
Coe, C. (1999). Psychosocial factors and psychoneuroimmunology within a lifespan perspective. In D. Keating and C. Hertzman (Eds.), Developmental health and the wealth of nations (pp. 201-219). New York: Guilford Press.
Cynader, M.S., and Frost, B.J. (1999). Mechanisms of brain development: Neuronal sculpting by the physical and social environment. In D. Keating and C. Hertzman (Eds.), Developmental health and the wealth of nations (pp. 153-184). New York: Guilford Press.
Davey Smith, G., Hart, C., Blane, D., Gillis, C., and Hawthorne, V. (1997). Lifetime socioeconomic position and mortality: Prospective observational study. British Medical Journal, 314, 547-552.
Davey Smith, G., Hart, C., Blane, D., and Hole, D. (1998). Adverse socioeconomic conditions in childhood and cause specific adult mortality: Prospective observational study. British Medical Journal, 316, 1631-1635.
Davis, R.J., and Collins, J.W. (1997). Differing birthweight among infants of US-born blacks, African-born blacks, and U.S.-born whites. New England Journal of Medicine, 337, 1209-1214.
Doll, R., and Peto, R. (1981). The causes of cancer. Oxford, England: Oxford University Press.
Drever, F., and Whitehead, M. (1997). Health inequalities. (DS #15, The Stationery Office). London: Office for National Statistics.
Duncan, G.J., and Brooks-Gunn, J. (Eds.). (1997). Consequences of growing up poor. New York: Russell Sage Foundation.
Duncan, G.J., and Brooks-Gunn, J. (2000). Family poverty, welfare reform, and child development. Child Development, 71, 188-196.
Duncan, G.J., Brooks-Gunn, J., and Klebanov, P.K. (1994). Economic deprivation and early childhood development. Child Development, 65, 296-318.
Eriksson, J.G., Forsen, T., Tuomilehto, J., Winter, P.D., Osmond, C., and Barker, D.J.P. (1999). Catch-up growth in childhood and death from coronary heart disease: Longitudinal study. British Medical Journal, 318, 427-431.
Evans, D.A., Beckett, L.A., Albert, M.S., Hebert, L.E., Scherr, P.A., Funkenstein, H.H., and Taylor, J.O. (1993). Level of education and change in cognitive function in a community population of older persons. Annals of Epidemiology, 3, 71-77.
Evans, R.G., Barer, M.L., and Marmor, T.R. (Eds.). (1994). Why are some people healthy and others not? The determinants of health of populations. New York: Aldine de Gruyter.
Fang, J., Madhavan, S., and Alderman, M.H. (1996, November). The association between birthplace and mortality from cardiovascular causes among black and white residents of New York City. New England Journal of Medicine, 335(21), 1545-1551.
Feldman, J.J., Makuc, D. M., Kleinman, J.C., and Cornoni-Huntley, J. (1989). National trends in educational differentials in mortality. American Journal of Epidemiology, 129(5), 919-933.
Francis, D., Diorio, J., Liu, D., and Meaney, M.J. (1999). Nongenomic transmission across generations of maternal behavior and stress responses in the rat. Science, 286, 1155-1158.
Frankel, S., Elwood, P., Sweetnam, P., Yarnell, J., and Davey Smith, G. (1996). Birthweight, body-mass index in middle age, and incident coronary heart disease. The Lancet, 348, 1478-1480.
Grotberg, E. (1995). A guide to promoting resilience in children: Strengthening the human spirit. The Hague, The Netherlands: Bernard van Leer Foundation.
Gunnar, M.R., and Barr, R. (1998). Stress, early brain development, and behavior. Infants and Young Children, 11, 1-14.
Guo, G., and Harris, K.M. (2000). The mechanisms mediating the effects of poverty on children’s intellectual development. Demography, 37, 431-447.
Hart, B., and Risley, T.R. (1995). Meaningful differences in the everyday experience of young American children. Baltimore: Paul H. Brookes.
Hertzman, C. (2000). The biological embedding of early experience and its effects on health in adulthood. Annals of the New York Academy of Sciences, 896, 85-95.
Hertzman, C., and Wiens, M. (1996). Child development and long-term outcomes: A population health perspective and summary of successful interventions. Social Sciences and Medicine, 43, 1083-1095.
Hertzman, C., Power, C., Matthews, S., and Manor, O. (2001). Using an interactive framework of society and lifecourse to explain self-rated health in early adulthood. Social Sciences and Medicine, 53, 1575-1585.
Hertzman, C., McLean, S., Kohen, D., Dunn, J., and Evans, T. (2002). Early development in Vancouver: Report of the Community Asset Mapping Project (CAMP). Available: http://www.earlylearning.ubc.ca [Accessed September 10, 2003].
Huxley, R.R., Shiell, A.W., and Law, C.M. (2000). The role of size at birth and postnatal catch-up growth in determining systolic blood pressure: A systematic review of the literature. Journal of Hypertension, 18, 815-831.
Keating, D., and Hertzman, C. (1999). Developmental health and the wealth of nations. New York: Guilford Press.
Kohen, D.E., Brooks-Gunn, J., Leventhal, T., and Hertzman, C. (2002). Neighbourhood income and physical and social disorder in Canada: Associations with young children’s competencies. Child Development, 73, 1844-1869.
Kuh, D.L., and Ben-Shlomo, Y. (1997). A life-course approach to chronic disease epidemiology: Tracing the origins of ill health from early to adult life. Oxford, England: Oxford University Press.
Kunst, A.E., Geurts, J.J.M., and van den Berg, J. (1995). International variation in socioeconomic inequalities in self-reported health. Journal of Epidemiology and Community Health, 49, 117-123.
Kunst, A.E., Groenhof, F., Mackenbach, J.P., and Health, E.W. (1998). Occupational class and cause specific mortality in middle aged men in 11 European countries: Comparison of population based studies. British Medical Journal, 316(7145), 1636-1642.
Lavis, J.N. (1997). An inquiry into the links between labour-market experiences and health. Doctoral dissertation, Harvard University.
Lynch, J.W., Kaplan, G.A., and Shema, S.J. (1997). Cumulative impact of sustained economic hardship on physical, cognitive, psychological, and social functioning. New England Journal of Medicine, 337(26), 1889-1895.
Marmot, M.G., and Wadsworth, M.E.J. (Eds.). (1997). Fetal and early childhood environment: Long-term health implications. British Medical Bulletin, 53, 3-9.
McDonough, P., Duncan, G.J., Williams, D., and House, J. (1997). Income dynamics and adult mortality in the United States, 1972 through 1989. American Journal of Public Health, 87, 1476-1483.
Miller, J., and Korenman, S. (1994). Poverty and children’s nutritional status in the United States. American Journal of Epidemiology, 140, 233-243.
Mustard, F., and Picherack, F. (2002). Early child development in British Columbia: Enabling communities. Toronto, Canada: Founders’ Network.
Neugebauer, R., Hoek, H.W., and Susser, E. (1999). Prenatal exposure to wartime famine and development of antisocial personality disorder in early adulthood. Journal of the American Medical Association, 282, 455-462.
Osmond, C., and Barker, D.J.P. (2000). Fetal, infant, and childhood growth are predictors of coronary heart disease, diabetes, and hypertension in adult men and women. Environmental Health Perspectives, 108(Suppl. 3), 545-553.
Pappas, G., Queen, S., Hadden, W., and Fisher, G. (1993). The increasing disparity in mortality between socio-economic groups in the United States, 1960 and 1986. New England Journal of Medicine, 329, 103-108.
Pollak, S., Cicchetti, D., and Klorman, R. (1998). Stress, memory, and emotion: Developmental considerations from the study of child maltreatment. Development and Psychopathology, 10, 811-828.
Post, R.M., and Weiss, S.R. (1997). Emergent properties of neural systems: How focal molecular neurobiological alterations can affect behavior. Development and Psychopathology, 9(4), 907-929.
Power, C., and Hertzman, C. (1997). Social and biological pathways linking early life and adult disease. British Medical Bulletin, 53(1), 210-221.
Power, C., Manor, O., and Fox, A.J. (1991). Health and class: The early years. London: Chapman Hall.
Power, C., Manor, O., and Matthews, S. (1999). The duration and timing of exposure: Effects of socio-economic environment on adult health. American Journal of Public Health, 89(7), 1059-1066.
Power, C., Li, L., and Manor, O. (2000). A prospective study of limiting longstanding illness in early adulthood. International Journal of Epidemiology, 29, 131-139.
Ravelli, A.C., Der Meulen, J.H., Osmond, C., Barker, D.J., and Bleker, O.P. (1999). Obesity at the age of fifty in men and women exposed to famine prenatally. American Journal of Clinical Nutrition, 70(5), 811-816.
Ravelli, G.P., Stein, Z.A., and Susser, M.W. (1976). Obesity in young men after famine exposure in utero and early infancy. New England Journal of Medicine, 295(7), 349-353.
Ross, D.P., and Roberts, P. (1999). Income and child well-being: A new perspective on the poverty debate. Ottawa, Canada: Canadian Council on Social Development.
Sapolsky, R.M. (1992). Stress, the aging brain, and the mechanisms of neuron death. Cambridge, MA: MIT Press.
Schweinhart, L.J., Barnes, H.V., and Weikart, D.P. (1993). Significant benefits: The High/ Scope Perry preschool study through age 27. Monographs of the High/Scope Educational Research Foundation, 10.
Seeman, M., and Lewis, S. (1995). Powerlessness, health and mortality: A longitudinal study of older men and mature women. Social Sciences and Medicine, 41, 517-525.
Seeman, T.E., and Crimmins, E. (2001). Social environment effects on health and aging. Annnals of the New York Academy of Sciences, 954, 88-117.
Seeman, T.E., and McEwan, B.S. (1996). Impact of social environment characteristics on neuroendocrine regulation. Psychosomatic Medicine, 58, 459-471.
Seeman, T.E., Berkman, L.F., Gulanski, B.I., Robbins, R.J., Greenspan, S.L., Charpentier, P.A., and Rowe, J.W. (1995). Self-esteem and neuroendocrine response to challenge: Macarthur studies of successful aging. Journal of Psychosomatic Research, 39, 69-84.
Susser, E., Neugebauer, R., Hoek, H.W., et al. (1996). Schizophrenia after prenatal famine: Further evidence. Archives of General Psychiatry, 53, 25-31.
Tremblay, R.E. (1999). When children’s development fails. In D. Keating and C. Hertzman (Eds.), Developmental health and the wealth of nations (pp. 55-71). New York: Guilford Press.
Tremblay, R.E., Masse, B., Perron, D., and LeBlanc, M. (1992). Disruptive behaviour, poor school achievement, delinquent behaviour, and delinquent personality: Longitudinal analyses. Journal of Consulting and Clinical Psychology, 60, 64-72.
Waters, M.C. (1999). Black identities: West Indian immigrant dreams and American realities. Cambridge, MA: Harvard University Press.
Werner, E.E. (1989). Children of the garden island. Scientific American, 260(4), 106-111.
Willms, D. (2000). Vulnerable children in Canada. Edmonton, Canada: University of Alberta Press.
Winkleby, M.A. (1992). Socioeconomic status and health: How education, income, and occupation contribute to risk factors for cardiovascular disease. American Journal of Public Health, 82, 816-820.




























