
5
Envisioning a Chemical Metaproteomics Capability for Biochemical Research and Diagnosis of Global Ocean Microbiomes
Mak A. Saito,a,* Chip Breier,bMike Jakuba,aMatthew McIlvin,aand Dawn Morana
INTRODUCTION
The oceans cover 70% of the Earth’s surface area and have an average depth of more than 4 km, representing the major component of the habitable environments on the planet. Thus, studying the microbiomes of the ocean represents a significant challenge due to the vastness and diversity of environments. These include free-living pelagic microbes, particle-associated microbial communities, sedimentary environments, and the remarkable microbial and metazoan communities found at the oases of hydrothermal vent communities. As economic activities have scaled to a global reach within the past two centuries, humans are increasingly impacting natural ecosystems, including the Earth’s critical life-sustaining biogeochemical cycles (Doney, 2010). These cycles encompass myriad elements and molecules within the geosphere, hydrosphere, and atmosphere domains of the Earth’s exposed environment (Falkowski et al., 2008). There is an expectation from society that scientists will identify and alert the public to environmental threats to allow policy makers to confront emerging threats. However, anthropogenic impacts have become global in extent, and often manifest themselves as multiple stressors occurring simultaneously. As a result, understanding the influence of multiple stressors on complex ecosystems presents a major scientific challenge (Gunderson et al., 2016). The development of new technologies has the potential to increase our ability to assess the impact of multiple environmental stressors; in particular, the advent of omics-based techniques has opened expansive observational windows into diversity and environmental function in manners previously not possible. In this chapter, we briefly discuss the development of chemical omic methodologies focused on the measurement of proteins within microbial populations throughout the oceans as a means to create a baseline of biochemical functions to detect changes in their activity.
The chemical omics fields, specifically proteomics and metabolomics, could have an important role in studies of oceanic microbes by allowing us to directly observe the molecules participating in biogeochemical reactions and the metabolic products. The oceanic biogeochemical cycles are typically influenced by a diverse assemblage of microbes conducting a similarly diverse array of biochemical reactions, which are integral to the elemental cycling of the oceans and the planet as a whole. In most cases, it appears that microbes are nutritionally limited in their chemical activities by specific chemical constraints, such as elements or molecules needed for constructing
___________________
a Woods Hole Oceanographic Institution.
b The University of Texas Rio Grande Valley.
* Corresponding Author: msaito@whoi.edu.
their biomass, or in the availability of substrates for key enzymes to act upon. The examples of this are numerous, where elements such as carbon, nitrogen, silicon, phosphorous, manganese, iron, cobalt, nickel, zinc, and cadmium have been studied for their potential nutritional control of microbial processes (Saito et al., 2008; Moore et al., 2013). Similarly, organic molecules used by heterotrophic respiration or inorganic substrates used in chemolithotropic reactions can limit the respiratory reactions of key microbial communities. Microbiologists have conducted pioneering studies to describe the diversity of the ocean’s communities, and we now know many of the important players in these ecosystems. Emerging chemical omic technologies that measure enzymes (proteomics) and metabolic products (metabolomics) have the potential to deepen our understanding of the biochemical functions, interactions, and controls of metabolic reactions. Such a functional analysis could complement biological diversity studies to create an invaluable baseline for the present microbial biogeochemistry of the oceans for comparison against changing ocean ecosystems.
Vitamin B12 as an Example of the Chemical Complexity Found in Ocean Microbiomes
An example of the complexity of chemical interactions in an oceanic microbiome comes from what appears to be the scarcest of all nutritional requirements: the vitamin B12 molecule, also known as cobalamin. B12 is the most structurally complex vitamin, containing a cobalt atom that is coordinated by a planar corrin ring and a dimethylbenzimidazole group. It requires more than 20 genes to biosynthesize, and is only made by some bacteria and Archaea (Rodionov et al., 2003). No eukaryote is known to have the complete B12 pathway, yet, in the oceans, roughly half of all phytoplankton require B12, while many others can utilize it to improve their metabolic efficiency (Droop, 2007; Helliwell et al., 2011). Key producers are the most abundant of those microbes, including the cyanobacteria (Prochlorococcus and Synechococcus) and the Thaumarchaeota (such as Nitrosopelagicus brevis) (Rodionov et al., 2003; Bonnet et al., 2010; Santoro et al., 2015). As a result, this complex biomolecule has a fascinating ecology in the microbial communities and microbiomes of the oceans. For example, in the tropical and subtropical oceans, it is produced by several major microbial groups, including cyanobacteria, likely as the pseudocobalamin variant chemical form; abundant tiny archaea; heterotrophic bacteria living on particles; and by abundant chemolithotrophic bacteria in the mesopelagic zone (Bonnet et al., 2010; Santoro et al., 2015; see Figure 5-1). Because of this abundance of cyanobacteria and microbial activity, there is likely a constant supply of cobalamin produced and released through the continual grazing by zooplankton and viral lysis of microbial B12 producers. Recent results have observed that pseudocobalamin is the chemical form of B12 produced by cyanobacteria, which implies this form may not be suitable for algae that live off the standard B12 chemical forms (Heal et al., 2014; Helliwell et al., 2016); however, pseudocobalamin could be subject to chemical reworking in bacterial salvage pathways (Escalante-Semerena et al., 1990).
In contrast to this model for B12 cycling in tropical and subtropical regions, a very different scenario appears to exist in the polar Ross Sea near Antarctica. The long and dark winter results in a large expanse of sea ice coverage around the Antarctic continent with minimal algal photosynthetic activity in the water column below; prokaryotic abundances are depressed during the ice-covered winter season due to a lack of dissolved organic matter production from phytoplankton (Ducklow et al., 2001). As a result, the prokaryotic sources of B12 to the polar ocean ecosystem become restricted throughout the winter (see Figure 5-1). As the sea ice retreats in the spring, those bacterial sources that rely on dissolved organic matter must wait for photosynthetic productivity to rebound. For unknown reasons, there are no appreciable marine cyanobacterial populations in the Southern Ocean that could supply B12. This scenario can explain the observations of B12–iron co-limitation of phytoplankton communities in the Ross Sea, where the addition of B12 with iron increases chlorophyll production significantly relative to iron-only controls (Bertrand et al., 2007).
Given that the Southern Ocean and Ross Sea are known to experience primary iron limitation, this secondary influence of B12 is likely ecologically important (Betrand et al., 2007, 2012; Sañudo-Wilhelmy, 2012). Notably, there are two dominant phytoplankton groups that comprise the annual blooms in the Ross Sea: diatoms and Phaeocystis antarctica (DiTullio and Smith, 1996). As the sea ice opens up in the Ross Sea polynya, the colony form of Phaeocystis blooms first in high abundances, which eventually yields to diatom populations as the bloom progresses. These Phaeocystis blooms are biogeochemically important, as they result in a significant and rapid export
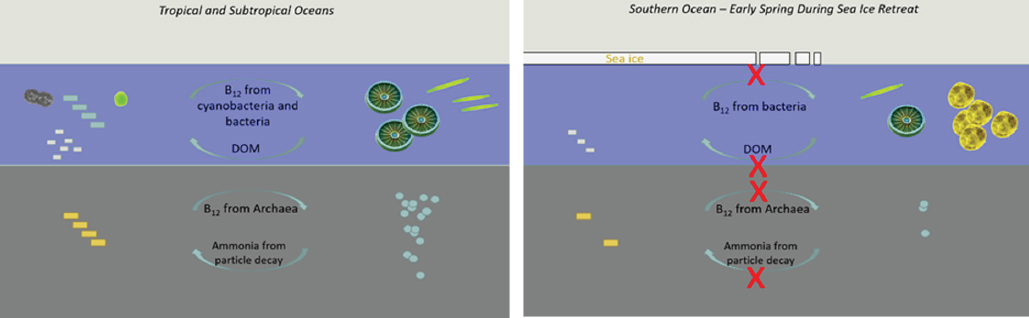
of carbon from the Ross Sea (DiTullio et al., 2000). There have been reports that deep, mixed layers and variable light could trigger the formation of colonies (Arrigo et al., 2010), and that the colonies could benefit Phaeocystis by allowing the storage of metals such as manganese (Davidson and Marchant, 1987). Recent metatranscriptomic and metaproteomic microbiome studies of the bacteria that live within the mucilage center of Phaeocystis colonies have identified bacteria capable of producing B12 in addition to B12 biosynthesis and iron storage proteins (Bertrand et al., 2015; Bender et al., in review). These findings imply that the microbiome of Phaeocystis colonies establish a chemical symbiosis, behaving like a vitamin bioreactor, where bacteria produce vitamin B12 in exchange for the dissolved organic matter from Phaeocystis. This scenario would give Phaeocystis an advantage relative to their diatom competitors and is consistent with incubation observations, where diatoms responded strongly to B12 and iron additions while Phaeocystis responded to iron, but not B12, additions (Bertrand et al., 2007). This example of interwoven chemical interactions between phytoplankton and prokaryotic communities provides an example of the tremendous complexity and diversity of oceanic microbiome systems that would be hard to emulate or predict from the study of simplified laboratory systems alone. The direct study of natural ecosystems and their chemical interactions will allow scientists a greater potential to grasp the full complexity of natural systems.
Metaproteomics and the Future Role for Chemical Omics in Understanding the Changing Oceans
In the past decade, mass spectrometry–based proteomic capabilities have improved tremendously, offering the promise of routine detection and quantitation of hundreds to thousands of proteins in any sample; a parallel set of advancements is occurring for metabolomics, which also includes the measurement of lipids (Soule et al., 2015). Together, these mass spectrometry–enabled omics could be considered chemical omics due to their focus on biomolecules, enzymes, and their metabolic products, in addition to the technical nature of the mass spectrometry instrumentation and chemical approaches required to measure these molecules. The application of proteomics to the oceans is complicated by specific challenges, particularly with regards to informatics, although species- and even ecotype- or subspecies-level resolution has been demonstrated by a new targeted metaproteomics method (Saito et al., 2015). An example of metaproteomics’ potential to observe microbiome chemical features is shown in Figure 5-2, which shows the distribution of the metalloenzyme nickel superoxide dismutase (NiSOD) across
SOURCE: From Saito, M. A., M. R. McIlvin, D. M. Moran, T. J. Goepfert, G. R. DiTullio, A. F. Post, and C. H. Lamborg. 2014. Multiple nutrient stresses at intersecting Pacific Ocean biomes detected by protein biomarkers. Science 345:1173-1177. Reprinted with permission from the American Association for the Advancement of Science.

NOTE: Hard hat provided for scale.
a 4,500-km stretch of the Central Pacific Ocean between Hawaii and Samoa (Saito et al., 2014). This NiSOD isoform appears to be highly abundant in this region of the Pacific Ocean for reasons that are not yet understood, but may be due to a need for increased superoxide dismutase activity as a result of intense iron scarcity present in this region and the resultant oxidative stress it induces. Moreover, the use of nickel in this enzyme may reflect a vestigial trait from the cyanobacteria’s evolution in anoxic oceans billions of years ago (Saito et al., 2003; Dupont et al., 2006). Finally, the NiSOD is only one of several thousand proteins identified in these types of datasets (see Figure 5-3) and, hence, reflects the tip of the iceberg in terms of making connections between cellular biochemistry, microbial ecology, and ocean biogeochemical cycling.
LEVERAGING ROBOTIC VEHICLE TECHNOLOGY: CLIO AS A GLOBAL OCEAN SAMPLING VEHICLE
The chemical approaches toward proteomics and metabolomics are powerful new tools that could be used to characterize the microbiomes of the oceans; yet, a major bottleneck in deploying these methods is the challenge of sampling the vastness of the global ocean environment. For the information-containing molecules DNA and RNA, there are methods that can exponentially amplify their concentration using polymerase enzymes, thereby increasing the signal intensity by many orders of magnitude from small samples. For the chemically functional proteins and their metabolite products, there is no comparable universal amplification approach. This difference is logical when considering the biology at hand: Nature uses only four base pairs each for DNA and RNA to enable facile enzymatically catalyzed reproduction. In contrast, nature uses more than 20 amino acids to generate the chemical functional diversity of protein polymers, thereby complicating any strategy to reproduce and amplify peptide polymers directly. The expectation that proteomics methods will mature to achieving sensitivity comparable to that of DNA naïvely overlooks these chemical differences between biomolecules. As a result, orders-of-magnitude larger sample sizes are needed to deeply characterize the proteome and metabolome of the ocean environments; yet, collecting the microbiome particles from 100 liters of seawater is logistically challenging and time consuming. For example, the ongoing GEOTRACES expeditions that collect full ocean depth and basin-scale chemical sections spend roughly half of their station time, or one-third of the entire expedition time, collecting microbial particles in seawater over the course of 50-70 days at sea. Efforts to sample the microbiome on a similar expedition using current technology would also require one-third of the expedition time.
In order to increase the efficiency of global-scale sampling of particulate material for chemical and biological analyses, it is becoming possible to take advantage of autonomous underwater vehicle (AUV) technology for high-resolution biochemical sampling; with funding from the National Science Foundation, we are building a new autonomous underwater vehicle that could contribute in this manner. Named Clio, this AUV was conceived to break the wire time sampling bottleneck by moving the particulate sampling off of the ship’s sampling wire onto an untethered vehicle, where simultaneous sampling of dissolved constituents and particulate biomass can occur, and effectively halving station sampling time (time required to acquire data at one position). The vehicle would be carried by a research vessel and deployed at each station, for example, on a transect across the Pacific Ocean. Clio would sample the water column while descending vertically to the ocean floor, and collect microbiome particulate samples at various depths while returning to the surface (see Figure 5-3); other water sampling operations would occur simultaneously on the ship’s hydrowire. In this manner, Clio could return hundreds of samples mapping the chemistry across the oceans (see Figure 5-2), but with more complete full ocean depth sampling and with much higher resolution. The use of Clio on future sectional expeditions could reduce station time by half, resulting in one-third shorter overall expedition length. The economic advantage to this mode of operation is significant: Clio would pay for itself in ship time savings with only a few expeditions. Clio will also improve the sample quality, as the in situ preservation of samples immediately after collection will preserve rapidly changing RNA and metabolites. The autonomous movement of the vehicle will enable high-resolution sampling through precise depth control, and allow feature-dependent opportunistic sampling based on sensor data while removing the influence of ship heave on sample collection depth.
A LOOK TO THE FUTURE: BUILDING THE PERSONAL MEDICINE RECORD FOR THE EARTH’S OCEANS
There are several areas that might benefit from the oceanic deployment of omics technologies. As mentioned in the introduction, the public has demonstrated a strong interest in learning the extent of anthropogenic impacts on the oceans. With microbial communities being the base of the ocean food webs, as well as the foundation of the Earth’s biogeochemical life support system, obtaining diagnostics about the stresses and changes in myriad microbial components is a worthy endeavor. This is particularly true as ecosystem and biogeochemical changes continue to occur throughout the global oceans. The chemical omics approaches of proteomics and metabolomics have the capacity to generate dense datasets that assess the diversity of chemical functions being deployed by major microbial constituents. The continued development of high-throughput AUV-assisted sampling, chemical extraction, mass spectrometry, and informatics will enable these chemical omics to extend their capabilities to study and diagnose entire ocean basins in the near future. Such a global deployment could create a much-needed baseline for ocean health prior to further anthropogenic alteration.
The omic approaches have, on occasion, been criticized as being correlative or nonmechanistic. Whether this is a fair criticism for all omic approaches is a debate beyond the scope of this chapter. Yet, it can be argued that this concern is less relevant for chemical omics, which are capable of measuring concentrations of enzymes, transporter proteins, and biochemical substrates and metabolites. Proteomics and metabolomics are similar to previously established techniques for protein abundance or metabolite abundance assays. The only differences are that biomolecules are being directly measured, typically with greater fidelity and sensitivity, and that instead of each assay only measuring one analyte, many analytes can now be measured simultaneously. As a result of this multianalyte ability, there is tremendous power in the chemical omics approach, albeit with a corresponding set of informatic challenges, particularly for complex environmental samples where many organisms are conducting a multitude of biochemical processes, often with significant biogeochemical impact.
Also, it is valuable to note that oceanography and, in particular, microbial biogeochemistry and the study of microbiome components are young fields; many major microbial taxa were only discovered within the past few decades. Prior assessments of environmental changes have had to rely on relatively simple bulk parameters such as chlorophyll and nutrient abundances over only a few decades. The billions of mass spectra that are being collected from chemical omics datasets could contribute to a deep record of what each of the major microbial taxa were experiencing in the environment at specific times and locations. Repeat analyses through seasons, years, and decades could offer scientists a deep understanding of how the major components of the oceans’ contribution to global biogeochemical cycles are currently operating and how they are changing with natural and human-induced perturbations. While we and others are currently attempting to prepare these analytical and sampling methods as rapidly as possible, we fear that our successors will still lament that we did not start soon enough prior to the multitude of measurable biogeochemical impacts on the oceans. Nevertheless, it seems clear that the path toward a sustainable coexistence of human economies with the Earth will inevitably require a deep understanding of the diversity and function of ocean ecosystems, and the chemical omics provide us with an unprecedented opportunity to greatly improve our understanding of the microbial chemical processes that sustain life in the oceans and on the Earth.
REFERENCES
Arrigo, K. R., M. M. Mills, L. R. Kropuenske, G. L. van Dijken, A.-C. Alderkamp, and D. H. Robinson. 2010. Photophysiology in two major Southern Ocean phytoplankton taxa: Photosynthesis and growth of Phaeocystis antarctica and Fragilariopsis cylindrus under different irradiance levels. Integr Comp Biol 50:950-966.
Bender, et al. In Review.
Bertrand, E. M., M. A. Saito, J. M. Rose, C. R. Riesselman, M. C. Lohan, A. E. Noble, P. A. Lee, and G. R. DiTullio. 2007. Vitamin B12 and iron co-limitation of phytoplankton growth in the Ross Sea. Limnol Oceanogr 52:1079-1093.
Bertrand, E. M., A. E. Allen, C. L. Dupont, T. Norden-Krichmar, J. Bai, and M. A. Saito. 2012. Influence of cobalamin starvation on diatom molecular physiology and the identification of a cobalamin acquisition protein. Proc Natl Acad Sci USA 109(26):E1762-E1771.
Bertrand, E. M., J. P. McCrow, A. Moustafa, H. Zheng, J. B. McQuaid, T. O. Delmont, A. F. Post, R. E. Sipler, J. L. Spackeen, and K. Xu. 2015. Phytoplankton–bacterial interactions mediate micronutrient colimitation at the coastal Antarctic Sea ice edge. Proc Natl Acad Sci USA 112:9938-9943.
Bonnet, S., E. A. Webb, C. Panzeca, D. M. Karl, D. G. Capone, and S. A. Sañudo Wilhelmy. 2010. Vitamin B12 excretion by cultures of the marine cyanobacteria Crocosphaera and Synechococcus. Limnol Oceanogr 55:1959-1964.
Davidson, A. T., and H. J. Marchant. 1987. Binding of manganese by Antarctic Phaeocystis pouchetti and the role of bacteria in its release. MarBiol 95:481-487.
DiTullio, G. R., and W. O. Smith, Jr. 1996. Spatial patterns in phytoplankton biomass and pigment distributions in the Ross Sea. J Geophys Res 101:18467-18477.
DiTullio, G., J. M. Grebmeier, K. R. Arrigo, M. P. Lizotte, D. H. Robinson, A. Leventer, J. P. Barry, M. L. VanWoert, and R. B. Dunbar. 2000. Rapid and early export of Phaeocystis antarctica blooms in the Ross Sea, Antarctica. Nature 404:595-598.
Doney, S. C. 2010. The growing human footprint on coastal and open-ocean biogeochemistry. Science 328:1512-1516.
Droop, M. R. 2007. Vitamins, phytoplankton and bacteria: Symbiosis or scavenging? J Phycol 29:107-113.
Ducklow, H., C. Carlson, M. Church, D. Kirchman, D. Smith, and G. Steward. 2001. The seasonal development of the bacterioplankton bloom in the Ross Sea, Antarctica, 1994-1997. Deep Sea Res II 48:4199-4221.
Dupont, C. L., S. Yang, B. Palenik, and P. E. Bourne. 2006. Modern proteomes contain putative imprints of ancient shifts in trace metal geochemistry. Proc Natl Acad Sci USA 103:17822-17827.
Escalante-Semerena, J., S. Suh, and J. Roth. 1990. cobA function is required for both de novo cobalamin biosynthesis and assimilation of exogenous corrinoids in Salmonella typhimurium. J Bacteriol 172:273-280.
Falkowski, P. G., T. Fenchel, and E. F. DeLong. 2008. The microbial engines that drive earth’s biogeochemical cycles. Science 320:1034-1039.
Gunderson, A. R., E. J. Armstrong, and J. H. Stillman. 2016. Multiple stressors in a changing world: The need for an improved perspective on physiological responses to the dynamic marine environment. Annu Rev Mar Sci 8:357-378.
Heal, K. R., L. T. Carlson, A. H. Devol, E. Armbrust, J. W. Moffett, D. A. Stahl, and A. E. Ingalls. 2014. Determination of four forms of vitamin B12 and other B vitamins in seawater by liquid chromatography/tandem mass spectrometry. Rapid Commun Mass Spectrom 28:2398-2404.
Helliwell, K. E., G. L. Wheeler, K. C. Leptos, R. E. Goldstein, and A. G. Smith. 2011. Insights into the evolution of vitamin B12 auxotrophy from sequenced algal genomes. Mol Biol Evol 28:2921-2933.
Helliwell, K. E., A. D. Lawrence, A. Holzer, U. J. Kudahl, S. Sasso, B. Kräutler, D. J. Scanlan, M. J. Warren, and A. G. Smith. 2016. Cyanobacteria and eukaryotic algae use different chemical variants of vitamin B12. Curr Biol 26:999-1008.
Moore, C. M., M. M. Mills, K. R. Arrigo, I. Berman-Frank, L. Bopp, P. W. Boyd, E. D. Galbraith, R. J. Geider, C. Guieu, S. L. Jaccard, T. D. Jickells, J. La Roche, T. M. Lenton, N. M. Mahowald, E. Maranon, I. Marinov, J. K. Moore, T. Nakatsuka, A. Oschlies, M. A. Saito, T. F. Thingstad, A. Tsuda, and O. Ulloa. 2013. Processes and patterns of oceanic nutrient limitation. Nat Geosci 6:701-710.
Rodionov, D. A., A. G. Vitreschak, A. A. Mironov, and M. S. Gelfand. 2003. Comparative genomics of the vitamin B12 metabolism and regulation in prokaryotes. J Biol Chem 278:41148-41159.
Saito, M. A., D. Sigman, and F. M. M. Morel. 2003. The bioinorganic chemistry of the ancient ocean: The co-evolution of Cyanobacteria and biogeochemical cycles at the Archean-Proterozoic boundary. Inorg Chim Acta 356:308-318.
Saito, M. A., T. J. Goepfert, and J. T. Ritt. 2008. Some thoughts on the concept of colimitation: Three definitions and the importance of bioavailability. Limnol Oceanogr 53:276-290.
Saito, M. A., M. R. McIlvin, D. M. Moran, T. J. Goepfert, G. R. DiTullio, A. F. Post, and C. H. Lamborg. 2014. Multiple nutrient stresses at intersecting Pacific Ocean biomes detected by protein biomarkers. Science 345:1173-1177.
Saito, M. A., A. Dorsk, A. F. Post, M. McIlvin, M. S. Rappé, G. DiTullio, and D. Moran. 2015. Needles in the blue sea: Subspecies specificity in targeted protein biomarker analyses within the vast oceanic microbial metaproteome. Proteomics 15:3521-3531.
Santoro, A. E., C. L. Dupont, R. A. Richter, M. T. Craig, P. Carini, M. R. McIlvin, Y. Yang, W. Orsi, D. Moran, and M. A. Saito. 2015. Genomic and proteomic characterization of “Candidatus Nitrosopelagicus brevis”: An ammonia-oxidizing archaeon from the open ocean. Proc Natl Acad Sci USA 112:1173-1178.
Sañudo-Wilhelmy, S. A., L.S. Cutter, R. Durazo, E. A. Smail, L. Gómez-Consarnau, E. A. Webb, M. G. Prokopenko, W. M. Berelson, and D. M. Karl. 2012. Multiple B-vitamin depletion in large areas of the coastal ocean. Proc Natl Acad Sci USA 109(35):14041-14045.
Soule, M. C. K., K. Longnecker, W. M. Johnson, and E. B. Kujawinski. 2015. Environmental metabolomics: Analytical strategies. Mar Chem 177:374-387.
This page intentionally left blank.








