
2
Overview and Assessment of the SterLim and JAXA Studies
The SterLim (Sterilisation Limits for Sample Return Planetary Protection Measures) report was presented to the committee at its meeting in 2017. An updated version of the SterLim report and the Japan Aerospace Exploration Agency (JAXA) report were presented at the committee’s meeting in 2018.1,2,3 The following sections are based on the review of the committee members for the various aspects of these reports and their findings. This chapter contains a brief outline of the work and the methodology the two teams followed. The structure of the chapter mirrors the process of microbial transfer, as modeled by the teams (Figure 2.1). This chapter also contains the committee’s review of the work presented to them for each step in the process.
The scenario that both teams considered for their respective studies assumed an initial microbial density in the surface of Mars and estimated how much of the microbial community would survive transfer from Mars to its moons. For the purposes of this report, the surface of Mars includes all material capable of being ejected from the planet during a major impact. Thus, the surface includes all material up to a depth of several km. Any martian microbes would pass through several phases of “sterilization” or dilution in their transfer from Mars to its moons. These phases are as follows:
- During the initial impact;
- During the aerodynamic heating when passing through the martian atmosphere;
- During the hypervelocity impact of the material on Phobos or Deimos and their subsequent recirculation and reimpact to the moons’ surface;
- From radiation on the surface of the moons; and
- From reformation and “gardening” of the moons’ surface from the natural meteoroid flux.
___________________
1 M. Patel, V. Pearson, D. Summers, D. Evans, A. Bennet, and P. Truscott, SterLim, 2017, “Sterilisation Limits for Sample Return Planetary Protection Measures ‘SterLim’: Presentation to the Committee on the Review of Planetary Protection Requirements for Sample Return from Phobos and Deimos,” http://sites.nationalacademies.org/cs/groups/ssbsite/documents/webpage/ssb_183902.pdf; referred to here as the “SterLim report.”
2 M. Patel, D. Summers, and D. Evans, 2018, “SterLim Team Presentation,” presentation to the committee on September 18.
3 A. Yamagishi and K. Fujita, 2018, “JAXA’s Planetary Protection Activities for Sample Return from Martian Moons,” TR 2018-13-02, presentation to the committee on September 18, 2018, http://sites.nationalacademies.org/SSB/CurrentProjects/SSB_181917.
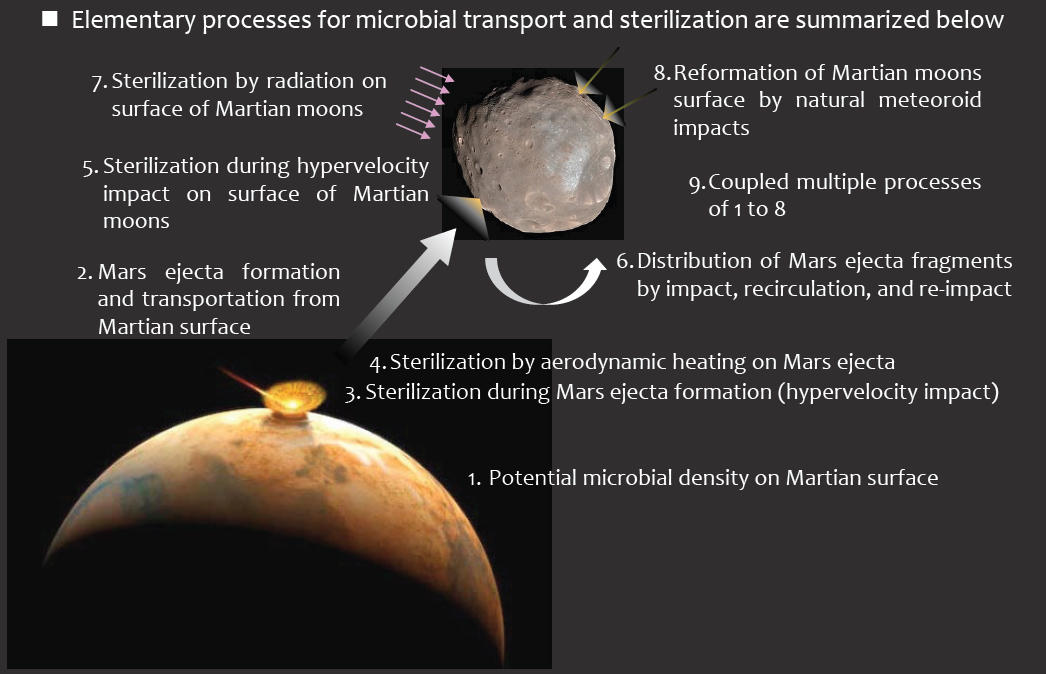
Each phase represents a specific model developed by the SterLim and JAXA teams or certain assumptions made. For example, Step 1 in Figure 2.1 requires that certain assumptions be made about the abundance of microbes in the martian surface material.
The subsequent sections of this chapter discuss each of the first eight steps outlined in Figure 2.1 and describe the modeling activities and assumptions made by both the SterLim and JAXA teams.
POTENTIAL MICROBIAL DENSITY ON THE MARTIAN SURFACE
The potential microbial density on Mars is, of course, a completely unknown parameter. The SterLim study considered the density to be similar to the Atacama Desert and used a range of possible bioloads (Table 2.1) to account for the uncertainties both in Earth measurements and in the potential variability of environments on Mars.
The JAXA report revised SterLim’s bioload estimates (see Table 2.1) based on measurements done in Antarctic analogues that are arid and cold (and thus, presumably, closer to a martian environment). The JAXA team concluded that Antarctic microbial density is of the same order or lower than the Atacama Desert. However, due to the lack of any scientific evidence on the baseline value for Mars, both studies have accepted a maximum value based on the Atacama studies, in order to provide a conservative assessment.
TABLE 2.1 Bioload Estimates Used by the SterLim and JAXA Teams Expressed in Colony Forming Units (cfu) per Kilogram of Martian Surface Material
| Martian Bioload Estimates Used | SterLim | JAXA |
|---|---|---|
| Low | 105 cfu/kg | Not used |
| Medium | 107 cfu/kg | Not used |
| High | 1010 cfu/kg | 108 cfu/kg |
NOTE: JAXA, Japan Aerospace Exploration Agency; SterLim, Sterilisation Limits for Sample Return Planetary Protection Measures.
Committee Assessment
Whether or not life exists on Mars is completely unknown. If it does exist, then its biochemical nature is even more of an unknown. If life actually exists on Mars, its molecular underpinning fits somewhere on a continuum with life as it exists on Earth—that is, carbon based, using water as a solvent, employing DNA to transfer genetic information, and employing RNA to control the synthesis of proteins—at one extreme, to organisms whose underlying chemistry is completely different from that of all known life on Earth. Planetary protection calculations are, in a sense, an exercise in due diligence, with researchers ruling out what they can based on what they know. If martian life is assumed to be fundamentally different from all life known on Earth, then researchers have no foundation on which to make planetary protection decisions. But, if researchers assume that life on Mars is basically similar to life on Earth, then they can make some progress. Mars today is cold and arid, so current understanding of life existing in the coldest and most arid regions on Earth may provide some insights into possible life-forms existing on Mars now and in the recent past.
Both the SterLim report and the JAXA report employed the values of the colony forming unit for the cell density of the Atacama Desert. However, the values of the unit are inappropriate as proxies of the cell density because colony forming ability generally depends on species of microbes. Namely, certain species of microbes have already been identified under the growth conditions on given medium agar plates, but others have not. The use of cell density values enumerated by direct cell count or biomolecules-based count (lipid, DNA, others) would have been more appropriate. Connon and coworkers reported that phospholipid fatty acid (PLFA) concentrations ranged from 2 × 105 to 7 × 106 cell equivalents per gram of soil, and Lester et al. reported that PLFA analysis indicated 2.0 × 106 to 1.0 × 107 cell equivalents per gram. Thus, the cell density of the Atacama Desert is likely to be greater than what was used by the SterLim and JAXA teams.4,5
Meanwhile, even if the dryness of Atacama is similar to Mars, it is probably true that the Atacama Desert is an energetically favorable environment for life compared with Mars. A close examination of the microbial communities in the soil of the Atacama Desert reveals that the ecosystem is governed by bacterial photosynthesis. In such ecosystems, several microbial species (cyanobacteria) are capable of using solar energy (a huge energy source) to fix carbon and thus form the basis of the food chain for the microbes surrounding the photosynthetic bacteria. Therefore, it would be better to use the values of cell density in the subsurface of the Atacama Desert. This would have minimized the effect of photosynthesis, which is an unlikely source of primary production on the martian surface given the strong ultraviolet flux.6 And it is probably reasonable, given that the cell density in the subsurface is much lower than that at the surface.
Regarding the Antarctic analogues, low microbial biomass was found by direct microscopic cell counts (1.4 to 5.7 × 106 cells per kg soil) in both the dry and ice-cemented permafrost. In this case, JAXA’s calculation (108 cells per kg) is sufficiently conservative.
___________________
4 S.A. Connon, E.D. Lester, H.S. Shafaat, D.C. Obenhuber, and A. Ponce, 2007, Bacterial diversity in hyperarid Atacama Desert soils, Journal of Geophysical Research 112:G04S17, doi:10.1029/2006JG000311.
5 E.D. Lester, M. Satomi, and A. Ponce, 2007, Microflora of extreme arid Atacama Desert soils, Soil Biology and Biochemistry 39:704-708.
6 T.C. Onstott, B.L. Ehlmann, H. Sapers, M. Coleman, M. Ivarsson, J.J. Marlow, A. Neubeck, and P. Niles, 2018, “Paleohosted Rock Life on Earth and the Search on Mars: A Review and Strategy for Exploration,” https://arxiv.org/abs/1809.08266.
MARS EJECTA FORMATION AND TRANSPORTATION FROM THE MARTIAN SURFACE
The ejection process following an impact on Mars was modeled by the SterLim team using the models developed by Melosh and coworkers in 2011 under a contract from NASA’s Office of Planetary Protection.7 The SterLim team did not consider any initial sterilization caused by the impact. Detailed modeling of Mars ejecta formation and subsequent transport from the martian surface was not part of the SterLim study’s statement of work. It was modeled only to the extent required to predict the mass transferred to the moons. The models were transformed to fit the needs of the Monte Carlo approach used by the SterLim team. They were normalized so that the total mass transported to Phobos in 10 million years corresponds to the value predicted by the models developed by Melosh et al. The SterLim models provided only the mass and velocity distributions of the martian ejecta.
The transit to the martian moons was modeled using orbital mechanics. The impact velocity distribution of martian ejecta on Phobos (or, by extension, on Deimos) obtained by the SterLim team corresponds well to the velocity distribution from Melosh’s models. In the Monte Carlo approach, impacts on Mars are modeled as stochastic events. This leads to a uniform transfer of mass from Mars to its moons throughout time.
In an addendum of the study, the SterLim team additionally looked at individual cratering events on Mars (“discrete ejections”) in general and focused on ejecta that would be generated from Zunil crater in particular. Zunil is a very young (~1-10 Ma), 10 km diameter impact crater on Mars, and is of particular interest to this study because materials from this impact event would by definition have had residence time <10 million years on the moons’ surfaces. In this addendum, the same approach was used to model the ejection process as for the original study.
The JAXA team considered five discrete events on Mars that formed craters that are >10 km in diameter and estimated to be younger than 10 million years. Five craters fit these criteria: Mojave, Tooting, McMurdo, Corinto, and Zunil. No other impact events on Mars were considered, although the contamination risk from unrecognized young craters was also investigated. From the dimensions of these five craters, possible combinations of impactor sizes, impact velocities, and impact angles were identified using a Monte Carlo approach and semi-empiric scaling laws. Corresponding numerical simulations were performed, and the ejecta generated by those simulated events propagated into Mars orbit and onto the moons. To reduce the calculation effort, five impact velocities (between 6 and 18 km/s) and six impact angles (between 0 and 75 degrees from the surface normal) were considered. For all craters, the actual location on Mars was included in the trajectory analysis. Specifically, for Zunil crater, the impact direction was restricted between northeast and east in accordance with a 2007 paper by Preblich et al.8
Both the SterLim and the JAXA teams reached the following conclusions:
- Large impacts (crater diameter >10 km) dominate the mass deposition on the martian moons.
- Ejecta deposited on the martian moons from craters older than 1 million years can be safely ignored due to the long residence of any microbes on the moon’s surfaces and the subsequent sterilization by the space radiation environment. (See “Sterilization by Radiation on Phobos/Deimos Surfaces,” later in this chapter.)
Both teams concluded that the last major impact on Mars with sufficient energy to deposit material on Phobos is the 10-km-diameter Zunil crater. Zunil is located in the equatorial Cerberus plains, one of the youngest areas on Mars and a potential target for future landed exploration.9 Thus, Zunil-derived materials will be present in the surface regolith of Phobos; less so for Deimos, which is more distant from Mars.
To estimate the mass of martian material ejected at speeds sufficient to reach Phobos’s orbit, numerical simulations were performed using the smoothed particle hydrodynamics (SPH) approach. The Tillotson equation of state, with a parameter set for granite, was used for both projectile and target. As neither a gravity nor a strength model was used in the numerical simulations, the results were considered dimensionless and thus scalable to any
___________________
7 H.J. Melosh, K.C. Howell, L. Chappaz, M. Vaquero, 2011, “Material Transfer from the Surface of Mars to Phobos and Deimos,” 22nd AAS/AIAA Space Flight Mechanics Meeting, Univelt Inc., San Diego, Calif.
8 B.S. Preblich, A.S. McEwen, and D.M. Studer, 2007, Mapping rays and secondary craters from the martian crater Zunil, Journal of Geophysical Research: Planets 112(E5), https://doi.org/10.1029/2006JE002817.
9 See, for example, A. McEwen, P. Lanagan, R. Beyer, L. Keszthelyi, and D. Burr, 2003, “Potential 2003 Landing Sites in the Cerberus Plains, Se Elysium Planitia,” presentation at the First Landing Site Workshop for MER 2003, https://www.lpi.usra.edu/meetings/mer2003/pdf/9022.pdf.
projectile size. The choice of granite, rather than the basalt likely to form much of the martian surface, was not explained. The simulations were validated by visual comparison to a single laboratory experiment with a 5 mm diameter polycarbonate projectile impacting a polycarbonate surface at 3.56 km/s with 45-degree impact incidence.
Committee Assessment
The models used by the two teams result in significantly different amounts of predicted mass ejected from Mars’s surface. This discrepancy is attributed to the different approaches followed by the two teams. The SterLim team relied on the work from Melosh and coworkers,10 who used semi-empirical impact crater models.11 This approach inherently includes most impact angles, but depends on the distribution of velocity of ejected fragment as a function of the mass of the ejected fragments, which is uncertain, particularly when including the impact angle.12 The JAXA team, in contrast, used the results from numerical simulations specifically performed for its study. These simulations predict that the mass ejected from a 45-degree impact is three to five times the mass ejected from a normal impact (depending on impact velocity),13 which is a significant increase. As a result, the mass transferred to Phobos is 1.12 × 106 kg according to Melosh et al. (2011) and 5 × 107 kg according to the JAXA team.14
The process of fragmentation and ejecta formation during hypervelocity impacts is currently not well understood. Specifically, the dependence of the mass of ejected material on impact angle is not described consistently in the literature. According to experiments, the final crater volume decreases with increasing impact angle (measured from the surface normal).15,16 In contrast, numerical simulations predict that the mass of ejected material increases with increasing impact angle.17,18 The results are not directly comparable. Experimental studies analyze the final crater volume (or, equivalently, ejecta mass), whereas numerical studies analyze the ejected mass above a threshold ejection velocity. Possible sources of this discrepancy are manifold, with two being the strength and porosity of the target material. These two latter factors were neglected in the JAXA simulations. It is known that the strength of the target material affects the final crater morphology of oblique impacts,19 which is also seen when comparing impact experiments on granite against experiments on low-strength materials.20,21 The influence of material strength on the ejecta formation process in general, and on the formation of fragments fast enough to reach the martian moons in particular, is not well known for these velocities and size scales. For fragments that are ejected at high velocities, and especially for the material transported to the martian moons, the influencing parameters, including material strength and porosity, are not fully constrained.
___________________
10 Melosh et al., 2011, “Material Transfer from the Surface of Mars to Phobos and Deimos.”
11 As described in, for example, J.E. Richardson, H.J. Melosh, C.M. Lisse, and B. Carcich, 2007, A ballistics analysis of the deep impact ejecta plume: Determining comet Temple 1’s gravity, mass, and density, Icarus 190:357-390, https://doi.org/10.1016/j.icarus.2007.08.001.
12 J.N. Head and H.J. Melosh, 2000, “Launch Velocity Distribution of the Martian Clan Meteorites,” Abstract 1937, Proceedings of the 31st Lunar and Planetary Science Conference, https://www.lpi.usra.edu/meetings/lpsc2000/pdf/1937.pdf.
13 See Figure 8-2 in K. Fujita, K. Kurosawa, H. Genda, R. Hyodo, T. Mikouchi, S. Matsuyama, and the Phobos/Deimos Microbial Assessment Team, 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons,” GNG-2018003.
14 K. Kurosawa, K. Fujita, H. Genda, R. Hyodo, T. Mikouchi, and Phobos/Deimos Microbial Contamination Assessment Team, 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons,” presentation to committee on September 19, http://sites.nationalacademies.org/cs/groups/ssbsite/documents/webpage/ssb_190400.pdf.
15 M.J. Burchell and L. Whitehorn, 2003, Oblique incidence hypervelocity impacts on rock, Monthly Notices of the Royal Astronomical Society 341(1):192-198, https://doi.org/10.1046/j.1365-8711.2003.06385.x.
16 T. Michikami, A. Hagermann, T. Morota, J. Haruyama, and S. Hasegawa, 2017, Oblique impact cratering experiments in brittle targets: Implications for elliptical craters on the Moon, Planetary and Space Science 135: 27-36, https://doi.org/10.1016/j.pss.2016.11.004.
17 Kurosawa et al., 2018, “Assessment of Phobos Microbial Contamination by Mars Ejecta.”
18 B. Artemieva and B. Ivanov, 2004, Launch of martian meteorites in oblique impacts, Icarus 171:84-101.
19 D. Elbeshausen, K. Wünnemann, and G.S. Collins, 2013, The transition from circular to elliptical impact craters, Journal of Geophysical Research: Planets 118(11):2295-2309, https://doi.org/10.1002/2013JE004477.
20 M.J. Burchell and L. Whitehorn, 2003, Oblique incidence hypervelocity impacts on rock, Monthly Notices of the Royal Astronomical Society 341(1):192-198, https://doi.org/10.1046/j.1365-8711.2003.06385.x.
21 T. Michikami, A. Hagermann, T. Morota, J. Haruyama, and S. Hasegawa, 2017, Oblique impact cratering experiments in brittle targets: Implications for elliptical craters on the Moon, Planetary and Space Science 135: 27-36, https://doi.org/10.1016/j.pss.2016.11.004.
Given the different approaches, the models of the SterLim and JAXA teams predict that significantly different masses of material are transported to the martian moons. For Phobos, the SterLim model predicts 1.6 × 106 kg (comparable to the 1.12 × 106 kg from Melosh et al.),22 while the JAXA model predicts 5 × 107 kg.23 The specific origin of this discrepancy between the two approaches could not be identified.
STERILIZATION DURING MARS EJECTA FORMATION
The SterLim team did not include any sterilization during Mars ejecta formation in its analysis because such investigations were not requested in its study’s statement of work. However, the JAXA team pointed out that considerable heat is generated by shock heating when an interplanetary impactor collides with Mars. The JAXA team performed a numerical simulation of a 45-degree impact at 3.5 km/s using the iSALE shock physics code.24,25,26 In this manner, it showed that ejecta faster than 3.8 km/s (i.e., that which can reach Phobos’s orbit) experience heating above 1000 K when considering internal friction and plastic deformation.27 Based on heat inactivation tests conducted by the SterLim team, its JAXA counterpart expected that this heating highly sterilizes the ejecta from Mars. In contrast to these impact physics models, the JAXA study did note that some experimental observations of martian meteorites do not show any signatures of shock heating. (See “Earth Inventory of Martian Meteorites,” in Chapter 1.) In summary, the JAXA team decided to assume a survival rate of 10 percent during Mars ejecta formation, but noted that this assumption may be too conservative.
Committee Assessment
The numerical simulation performed by the JAXA team suggests that the ejecta accelerated sufficiently to reach the martian moons experience significant heating that enables sterilization.28 Additionally, the JAXA team reviewed investigations of martian meteorites, some of which show significant shock heating and others showing no signs of shock heating. (See “Earth Inventory of Martian Meteorites,” in Chapter 1.) The JAXA team was therefore not confident in the validity of its results. The simulation was performed for a comparatively low impact speed of 3.5 km/s. At such a low speed, the material accelerated to ejection velocity most likely originates from near the contact surface, where plastic deformation (and heating) is most significant. For higher impact speeds, material farther from the contact surface can also reach ejection velocity, presumably with much smaller plastic deformation. Also, shock heating is a highly localized process, and numerical simulations addressing it require very high spatial resolution. The committee was unable to define a survival rate based on the information available. However, the proposed survival rate of 10 percent is a reasonable estimate, albeit one lacking significant experimental evidence.
___________________
22 Melosh et al., 2011, “Material Transfer from the Surface of Mars to Phobos and Deimos.”
23 Kurosawa et al., 2018, “Assessment of Phobos Microbial Contamination by Mars Ejecta.”
24 A.A. Amsden, H.M. Ruppel, and C.W. Hirt, 1980, “SALE: A Simplified ALE Computer Program for Fluid Flow at All Speeds,” LA-8095, https://inis.iaea.org/collection/NCLCollectionStore/_Public/11/571/11571883.pdf.
25 B.A. Ivanov, S. Deniem, and G. Neukum, 1997, Implementation of dynamic strength models into 2-D hydrocodes: Applications for atmospheric breakup and impact cratering, International Journal of Impact Engineering 20:411.
26 K. Wünnemann, G.S. Collins, and H.J. Melosh, 2006, A strain-based porosity model for use in hydrocode simulations of impacts and implications for transient crater growth in porous targets, Icarus 180(2):514-527, https://doi.org/10.1016/j.icarus.2005.10.013.
27 See Figure 4-1b of K. Kurosawa and H. Genda, 2018, Effects of friction and plastic deformation in shock-comminuted damaged rocks on impact heating, Geophysical Research Letters 45(2):620-626, https://doi.org/10.1002/2017GL076285.
28 K. Kurosawa and H. Genda, 2018, Effects of friction and plastic deformation in shock-comminuted damaged rocks on impact heating, Geophysical Research Letters 45(2):620-626, https://doi.org/10.1002/2017GL076285.
STERILIZATION BY AERODYNAMIC HEATING OF MARS EJECTA
The passage through the martian atmosphere was considered only by the JAXA team and not by the SterLim team. Using computational fluid dynamics simulations, heating and aerodynamic deceleration of single spherical ejecta fragments during the passage through the martian atmosphere were assessed. For this step in the chain, those parts of the fragment were considered sterilized where a temperature above 773 K was maintained for more than 0.5 s.29 The JAXA team concluded that aerodynamic heating could cause a microbial survival rate below 14 percent for 2 cm size fragments depending on impact angle. For fragments above 4 cm size, more than 50 percent of microbes were predicted to survive. Therefore, sterilization by aerodynamic heating was not considered to be significant in the further analyses because particles less than 10 cm are unlikely to reach Mars orbit due to aerodynamic deceleration.
Committee Assessment
The JAXA model considers only single ejecta fragments passing through the martian atmosphere. The major conclusion from the aerodynamic analysis was that particles smaller than 10 cm are unlikely to reach Mars orbit due to aerodynamic deceleration. However, an impact event creates numerous particles that pass through the atmosphere within a very short time period. Therefore, it is to be expected that much of the ejecta will interact with a disturbed atmosphere.
A mass comparison shows that the ejected mass traveling at high velocities is approximately of the same order of magnitude as the disturbed atmospheric mass. According to McEwen et al.,30 numerical simulations of the Zunil impact event suggest that 1.5 km3 of martian rocks were ejected at velocities greater than 1 km/s, which translates to 4 × 109 kg assuming a density of 2.6 g/cm3. The mass of the atmosphere directly above the crater is roughly 1.3 × 1010 kg (calculated from 600 Pa Mars surface pressure, 3.7 ms–2 Mars surface gravity, and 10.1 km crater diameter). Hence, the influence of the ejecta particles on the atmosphere is considered significant. This effect reduces the aerodynamic deceleration and brings into question any subsequent analyses making use of this lower limit on the size of particles capable of escaping the martian atmosphere. However, it is clear, as the JAXA team concluded, that aerodynamic heating of ejecta during passage through the martian atmosphere does not cause any significant sterilization.
STERILIZATION DURING HYPERVELOCITY IMPACT ON PHOBOS/DEIMOS SURFACES
Hypervelocity impact experiments, designed to assess impact survivability for four types of organisms, were carried out by the SterLim team using the Open University’s two-stage, light-gas gun. In each experiment, selected organisms (see “Sterilization by Radiation on Phobos/Deimos Surfaces,” later in this chapter, for details) were loaded into a hole drilled into the rear face (relative to the velocity vector) of cylindrical projectiles. Experimental design was confined to one projectile material (solid basalt slugs, 3 mm in diameter). The one target material was a Phobos regolith simulant sieved to particle sizes 400 μm and under, with an approximate porosity of 23 percent and bulk density of 1.51 g/cm3. The target buckets were small, only 70 mm in diameter,31 to reduce the required regolith mass to ~100 g per experiment. Exploration of sensitivity to target and projectile material choices was not pursued apart from early attempts with concrete targets, which sterilized everything.
The results presented for organism survival at velocities 0.5-1.8 km/s exhibited many orders of magnitude of spread at a given velocity. The SterLim team assessed that no organisms will be killed as a result of an impact at 0.5 km/s, attributing losses at those speeds to flawed collection methods. In a given experiment, the fraction of successfully recovered projectile fragments was very difficult to assess, given compositional similarity between the target and projectile materials chosen and insufficient target confinement. At larger impact speeds, the SterLim team
___________________
29 See Section 5.4 of Fujita et al., 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons.”
30 A.S. McEwen, B.S. Preblich, E.P. Turtle, N.A. Artemieva, M.P. Golombek, M. Hurst, R.L. Kirk, D.M. Burr, P.R. Christensen, 2005, The rayed crater Zunil and interpretations of small impact craters on Mars, Icarus 176(2):351-381, https://doi.org/10.1016/j.icarus.2005.02.009.
31 See Figures 8 and 9 of Patel et al., 2018, “SterLim Team Presentation.”
asserted that the collection methods were more robust and projectile/organism material was not lost. Experiments were carried out only up to 1.8 km/s, while the stated requirements for the SterLim study included velocities up to 4.5 km/s.32 This requirement was based on the expected velocity distribution of martian material impactors on Phobos and Deimos. The primary diagnostic used for the experiments was the unsterilized fraction of the organisms; planned pyrometer measurements were not successful.
Projectile survival is known to be highly dependent upon impact angle, which affects the peak pressures experienced and the way in which the impactor fails.33 The SterLim study included vertical shots (90 degrees to surface) and near-horizontal shots with a tilted target bucket (~50 degrees to surface).
The JAXA team performed Monte Carlo simulations to obtain the distribution of collision velocity of Mars rocks onto Phobos and Deimos. It showed that most of the primary ejecta has an impact velocity between ~1-to-20 km/s. The JAXA team used the sterilization data obtained by the SterLim team as input directly into its calculations and did not conduct any additional hypervelocity impact experiments. Unlike the SterLim team, it fit the SterLim sterilization versus impact velocity data directly (without any modeling) using a specific exponential function that it assumed could be applied at the high velocities of the Mars rock impact on the moons.34
Committee Assessment
Several issues that were not considered during the SterLim impact experiments and their subsequent analyses may have had significant influence on the results. Since the SterLim results were then used directly by the JAXA team, those errors and uncertainties propagated into the JAXA models. In addition, the JAXA team drew some conclusions from the SterLim results that were very broad generalizations in terms of sterilization from hypervelocity impact, and the committee heard explicitly from the SterLim team that it believed that those generalizations were unfounded.
In each of the SterLim impact experiments, organisms were loaded into a hole drilled into the rear face of cylindrical projectiles. This raised the possibility that high gas temperatures during the launch phase may have contributed to organism sterilization prior to impact because the sabot, into which the projectile is loaded, did not seal off the gas from the projectile. An estimation of the driver gas conditions during launch yields temperatures in excess of 800 K during launch to 2 km/s, which can last for up to 0.8 ms (assuming a 0.1 g projectile, a 0.8 m launch tube, and 1 MPa initial nitrogen fill pressure).35,36 At lower velocities, the temperatures are considerably lower, but last longer (360 K for 3 ms for a 500 m/s launch, 540 K for 1.6 ms for a 1 km/s launch).
Additionally, as the shock wave hits the rear face of the projectile during impact, the projectile will go into tension, causing fragments to be launched at speeds up to twice the particle velocity in the reference frame of the projectile. Hence, the backside of the projectile is a particularly unfavorable location for organism placement, because of likely loss of biological material, even though it may be perhaps the most favorable location for survival in the laboratory experiment because organisms are separated from the projectile before heat is effectively transferred to the organisms. Thus, it is not clear that the SterLim team’s assertion that projectile/organism material was not lost during impact is well supported.
The SterLim team justified ending experiments at 1.8 km/s due to high levels of organism sterilization at that velocity. However, given the large variations in sterilization results, this is not a particularly robust argument. Data at higher velocities would have been useful for validation of the complementary numerical simulations, provided that appropriate diagnostics were employed. Such data would also have been useful in providing better guidance to the JAXA team in terms of sterilization due to hypervelocity impact.
___________________
32 See Requirement 190 in Patel et al., 2018, “SterLim Team Presentation.”
33 See, for example, R.T. Daly and P.H. Schultz, 2016, Delivering a projectile component to the vestan regolith, Icarus 264:9-19.
34 See Equation 6.1 in Fujita et al., 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons.”
35 J. Lukasiewicz, 1967, Constant acceleration flows and applications to high-speed guns, AIAA Journal 5(11):1955-1963, https://doi.org/10.2514/3.4346.
36 G.P.T. Wilenius, P.L. Cowan, and M. Cloutier, 1964, “The Constant Base Pressure Light Gas Gun,” in Proceedings of the 3rd Hypervelocity Techniques Symposium, Denver Research Institute, Denver, Colo.
The SterLim team claimed that impact angle effects were unimportant, due to no discernible trend between the 90- and 50-degree shots. However, since the variability in the data was very high, even within each impact angle group, this claim was not well supported. Again, problems in experimental design and collection methods cast doubt on the conclusion about impact angle. As the impact angle probability distribution is sinusoidal and centered around 45 degrees for natural planetary impacts, near-vertical impacts are exceedingly rare.37,38
Understanding the role of impact angle in organism survival is an important piece of the problem. The SterLim team actually illustrated this point, to some extent, with a handful of three-dimensional (3D) simulations presented in the numerical impact study: A significantly larger fraction of the projectile was not heated to the sterilization limits in the 30-, 20-, and 10-degree simulations, as compared with a two-dimensional (2D), 90-degree impact simulation.
Throughout the study, it was claimed that a regolith “bounce back” effect would result in nearly all projectile material remaining at the surface. There is no physical justification for this assertion, however. The SterLim team set the depth deposited to be one radius of the impactors in its modeling, although experimentalists recognize that fragments can be buried to depths of more than one impactor radius in hypervelocity impact experiments using porous particulate targets (regolith-like).39 Depth of burial for projectile material is a significant factor to consider, as it could preserve organisms from sterilizing radiation on the surface of Phobos or Deimos.
Overall, the experimental hypervelocity impact data generated during the SterLim study is limited with respect to the large spectrum of possible impact conditions on martian moons, could be biased, and is not conclusive. Given the small footprint of the data within the vast parameter space, extrapolations drawn from this data seemed to the committee to be ill-advised. The committee notes that SterLim’s impact data was then used to calibrate the exponential function that the JAXA group used to estimate the likely sterilization due to impact.40
DISTRIBUTION OF MARS EJECTA FRAGMENTS BY IMPACTS, RECIRCULATION, AND REIMPACT
The SterLim team assumed that spatially uniform martian ejecta impact with the moons with a random impact angle.41 Part of the martian materials is deposited on the surface of the moons, and the rest is ejected from the satellites. A part of the ejected materials becomes a “debris cloud” orbiting Mars. Based on the numerical simulation of impact,42 it was assumed that the impacting material with angle between vertical (0 degree from the surface normal) and 45 degrees is fully deposited on the moons. It was also assumed that the deposited fraction linearly decreases with the angle for highly oblique incidence (>45 degrees). As a result, the percentage deposited on the Phobos surface at the first collision will be 78 percent and on the Deimos surface will be 82 percent. The burial depth of the martian material left on the surface depends on the properties of the moons’ regolith and the size of the impactor. Burial depths of one to a few times the radius of the impactor—that is, fragments of martian ejecta—were expected, and the SterLim team reported burial depths of one ejecta-fragment radius in its report. Most of the martian materials were expected to be deposited near the surface.
The SterLim team assumed that the normal velocity component of the ejecta from the moon is 40 percent of the normal component of the impact velocity and pointing away from the moon. The tangential component
___________________
37 G.K. Gilbert, The Moon’s Face: A Study of the Origin of its Features, 1893 presidential address to the Philosophical Society of Washington and reported in Science 21(539):305-307, http://science.sciencemag.org/content/sci/ns-21/539/305.4.full.pdf.
38 E.M. Shoemaker, 1962, “Interpretation of Lunar Craters,” pp. 283-359 in Physics and Astronomy of the Moon (Z. Kopal, ed.), Academic Press, London.
39 See Section 7.2.4 in Thales Alenia Space, 2018, “Evaluation of the Level of Assurance that No Unsterilized Martian Material Naturally Transferred to Phobos (and Deimos) Is Accessible to a Phobos (and Deimos) Sample Return Mission,” SterLim-Ph2-TAS-TN21, June 9, http://sites.nationalacademies.org/cs/groups/ssbsite/documents/webpage/ssb_190403.pdf; referred to as “SterLim TN-21.”
40 See Equation 6-1 in Fujita et al., 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons.”
41 See Thales Alenia Space, 2018, “Sterilisation Limits for Sample Return Planetary Protection Measures: TN 19 Description of the Sterilisation Statistical Model,” SterLim-Ph2-TAS-TN19, June 9, http://sites.nationalacademies.org/cs/groups/ssbsite/documents/webpage/ssb_190404.pdf, and SterLim TN-21.
42 See Fluid Gravity Engineering Ltd., 2017, “Report on the Hypervelocity Impact Modelling,” SterLim-PH2-FGE-TN-18, May 10, http://sites.nationalacademies.org/SSB/CurrentProjects/SSB_181917#presentations.
was assumed to be from 76 percent (45-degree impact to the surface) to 100 percent (grazing impact). The team considered that the material going into the cloud undergoes little impact heating during impact and excavation, and so will not be sterilized. It assumed that the ejected material remains at the original size, and therefore the orbital motion is hardly affected by solar irradiation. Most of the ejecta escaped from the Mars system, but half of the remaining (i.e., cloud component) reimpacted with the moons and the other half impacted with Mars. The team assumed that the radiation sterilization is negligible for the cloud component. It showed that the reimpacting velocity distribution to Phobos/Deimos shifts to a lower velocity than the velocity distribution of the direct component from Mars, and sterilization of the cloud component due to impact heating during the second collision is less likely to occur than direct impact component—that is, those deposited at the first collision.
As previously described (see “Mars Ejecta Formation and Transport from the Martian Surface,” earlier in this chapter), the JAXA team estimated the amount of martian ejecta with a velocity that could reach the moons by SPH simulations, by changing the impactor’s angle and velocity toward Mars.43 Based on this SPH modeling, the JAXA team estimated the impact velocity and angle of the Mars ejecta onto Phobos and Deimos by the Monte Carlo method. Since the size distribution of Mars ejecta colliding with the moons is unknown, the number of martian rocks colliding at Phobos and Deimos was estimated from the total mass of the ejecta while assuming that the martian rocks were all 10 cm diameter spheres.
The JAXA team estimated the diameter of the “Mars-rock-crater” on the moons using pi-group scaling.44 Since the stress experienced by martian rocks when impacting onto the regolith of the moon’s surface exceeds the estimated compressive strength of the rocks, the team expected that martian rocks will be broken into pieces at the time of impact to form a lens-shape region at the bottom of the resulting crater.45 A significant fraction of the impactor fragments (i.e., martian rock fragments) is ejected from the regolith surface depending on the collision angle and velocity of martian rocks to the moon’s surface.46 In the case of ejecta from Zunil crater, the percentage of impactor fragments remaining in craters was estimated as 22 and 29 percent for Phobos and Deimos, respectively. A 10 cm diameter martian rock forms a crater with a diameter of ~10 m and a lens thickness of 1 m.
Part of the ejected fragments from the moon’s surface will again fall on the surface of the moons. The JAXA team reported that the average thickness of the reaccumulated material will be 30 microns for Phobos and 1 micron for Deimos as a result of the Zunil impact, for example. The team assumed that the thickness of the reaccumulated layer (“global thin layer”) is 0.1 mm and expected that it is quickly sterilized by radiation.
Committee Assessment
The SterLim and JAXA teams obtained results for the fraction of martian ejecta deposited on Phobos or Deimos. SterLim expected that about 80 percent is deposited at the first impact to the moons, whereas the JAXA team expected that about 70-80 percent is ejected. The estimations of the two teams are based on the different and limited experimental data (the SterLim team used its own data, and the JAXA team used literature data). Therefore, a factor of uncertainty remains in the fraction deposited at the first impact.
To be conservative, the depth of deposition (one radius of Mars rocks) assumed by the SterLim team needs a modification. The depth of deposition in the JAXA team’s model is conservative. The JAXA team’s calculations showed that some 80 percent of the fragments of Mars rocks are distributed in a global layer some 0.1 mm thick. Whereas, the remaining 20 percent is mixed with regolith and confined to 1 m thick layers (“collapsed lens”) in the bottoms of the craters formed by the impacting martian material.
The model of the JAXA team assumes severe comminution of impactors during the penetration or the ejection. It is true that the dynamic ram pressure exceeds the compressive strength of intact basaltic rocks and Mars rocks would be fragmented. However, this does not mean that the rocks would be fully pulverized into dust (<0.1 mm), but possibly into mm or larger size. Laboratory experiments have shown that the degree of fragmentation depends
___________________
43 See Section 8 of Fujita et al., 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons.”
44 Ibid.
45 See the “collapsed lens” in Figure 9-2 of Fujita et al., 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons.”
46 R.T. Daly and P.H. Schultz, 2016, Delivering a projectile component to the vestan regolith, Icarus 264:9-19.
on the ratio of the compressive (or tensile) strength of rocks and the applied stress.47,48 The size of the largest fragment is highly dependent on the size of the Mars rock and the strength of the Mars rocks that are damaged during their acceleration as they are ejected from the surface of Mars.
STERILIZATION BY RADIATION ON PHOBOS/DEIMOS SURFACES
The SterLim report investigated the killing effects of protons, alpha particles, and gamma rays on test organisms. Ultraviolet light (e.g., UVC) and particle irradiations (e.g., charged particles such as protons and alpha particles) were not factored in microbial survival on Phobos and Deimos, because these forms of radiation are blocked by the top few millimeters of the moons’ regolith. These organisms included the extremely radiation resistant bacterium Deinococcus radiodurans, the biomedical indicator strains Brevundimonas diminuta and Bacillus atrophaeus, and an ssRNA virus MS2 coliphage. The representative organisms were irradiated with the objective of determining the radiation conditions required to ensure that sterilization limits are achieved.
Anaerobic, desiccated, and deeply frozen preconditions for the survival of life on Mars before transfer to Phobos and Deimos can be simulated in the laboratory. It follows that the SterLim team was tasked to simulate and demonstrate the level of biological inactivation of materials transferred from Mars to Phobos and Deimos. In an attempt to model better the effects of radiation on possible extant life forms on the surfaces of Mars and its moons, the effects of ionizing radiation (gamma radiation) were studied on desiccated cells and viruses under deeply frozen anaerobic conditions.
Specifically, the SterLim team applied a fixed number of bacteria and viruses directly onto aluminum slides, which were dried, frozen (193 K) and stored under nitrogen gas, and then exposed to acute doses (0-50 kGy) of gamma radiation. Finally, colony- and plaque-forming unit assays for bacteria and viruses, respectively, were used to gauge survival on the irradiated slides. Special emphasis was placed on D. radiodurans, the most characterized model for extreme radiation resistance.49,50,51D. radiodurans is capable of surviving 16 kGy under aqueous conditions, 10-times greater than baker’s yeast (Saccharomyces cerevisiae), 20-times greater than Escherichia coli bacteria, and 3,000-times greater than human cells.52 In comparison, viruses are generally the life-form most resistant to radiation.53 Microorganisms were held in their original growth medium, then transferred to the surfaces of sterilized slides, then dried under ambient atmospheric conditions, and finally irradiated.
The results of the irradiation experiments undertaken by the SterLim team are summarized as follows: For the test organisms, the survival metrics of bacteria and viruses exposed to ionizing radiation in the form of gamma rays under desiccated and deeply frozen conditions were slightly greater, both in form and scale, to survival metrics reported by others for gamma irradiations performed under aqueous conditions at 273 K.54
___________________
47 Y. Takagi, H. Mizutani, and S.-I. Kawakami, 1984, Impact fragmentation experiments of basalts and pyrophyllites, Icarus 59:462-477.
48 H. Nagaoka, S. Takasawa, A.M. Nakamura, and K. Sangen, 2013, Degree of impactor fragmentation under collision with a regolith surface—Laboratory impact experiments of rock projectiles, Meteoritics and Planetary Science 49:69-79.
49 M.J. Daly, L. Ouyang, P. Fuchs, and K.W. Minton, 1994, In vivo damage and recA-dependent repair of plasmid and chromosomal DNA in the radio-resistant bacterium Deinococcus radiodurans, Journal of Bacteriology 176:3508-3517.
50 K.S. Makarova, L. Aravind, Y.I. Wolf, R.L. Tatusov, K.W. Minton, E.V. Koonin, and M.J. Daly, 2001, Genome of the extremely radiation resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics, Microbiology and Molecular Biology Reviews 65:44-79.
51 M.J. Daly, E.K. Gaidamakova, V.Y. Matrosova, A. Vasilenko, M. Zhai, A. Venkateswaran, M. Hess, M.V. Omelchenko, H.M. Kostandarithes, K.S. Makarova, L.P. Wackett, J.K. Fredrickson, and D. Ghosal, 2004, Accumulation of Mn(II) in Deinococcus radiodurans facilitates gamma-radiation resistance, Science 306:1025-1028.
52 A. Sharma, E.K. Gaidamakova, O.E. Grichenko, V.Y. Matrosova, V. Hoeke, P. Klimenkova, I.H. Conze, et al., 2017, Across the tree of life, radiation resistance is governed by antioxidant Mn2+, gauged by paramagnetic resonance, Proceedings of the National Academy of Sciences U.S.A. 114:E9253-E9260.
53 E.K. Gaidamakova, I.A. Myles, D.P. McDaniel, C.J. Fowler, P.A. Valdez, S. Naik, M. Gayen, P. Gupta, A. Sharma, P.J. Glass, R.K. Maheshwari, S.K. Datta, and M.J. Daly, 2012, Preserving immunogenicity of lethally irradiated viral and bacterial vaccine epitopes using a radio-protective Mn(2+)-peptide complex from Deinococcus, Cell Host Microbe 12(1):117-124.
54 A. Sharma, E.K. Gaidamakova, O.E. Grichenko, V.Y. Matrosova, V. Hoeke, P. Klimenkova, I.H. Conze, et al., 2017, Across the tree of life, radiation resistance is governed by antioxidant Mn2+, gauged by paramagnetic resonance, Proceedings of the National Academy of Sciences U.S.A. 114:E9253-E9260.
The JAXA report used the SterLim data for its analysis, averaging the survival rate for specific depth, in order to facilitate the modeling of sample collection with a coring approach, as planned by MMX (see Figure 1.4).
Committee Assessment
The SterLim team’s irradiation setup included placing samples of bacteria and viruses directly onto the metal surfaces of aluminum strips. However, aluminum exposed to gamma radiation causes backscattering of electrons at surfaces and forms reactive oxides.55,56 This increases the proximal dose rates and damage to cells and viruses in contact with the metal during irradiation. The radiation resistance of desiccated Deinococcus bacteria would be expected to be higher if irradiated on a chemically inert surface such as polystyrene. Moreover, aluminum metal-cell interactions are not anticipated on the surfaces of Mars or its moons.57 Overall, the use of aluminum as a simulant environment for irradiation on Mars/Phobos is problematic.
Published desiccation protocols for bacteria including Deinococcus spp. involve transferring cells to inert surfaces, then drying in sealed chambers containing desiccants (e.g., Drierite). Polystyrene surfaces prevent secondary reactions with biosamples, and Drierite accelerates evaporation, lessening cell growth during the desiccation process itself, which typically lasts one week.58 In the absence of hermetically sealed desiccation chambers, the SterLim bacteria might have been washed and resuspended in phosphate buffer prior to desiccation—to prevent continued growth, which can cause large standard deviations in desiccation survival curves.59
In subsequent discussions, the committee refers to the higher gamma radiation survival values reported for deinococcal cells irradiated as deeply frozen aqueous preparations.
The Radiation Environment on the Martian Moons
A critical issue upon which support or not for the presence of preserved life on Phobos and Deimos transferred from Mars is radiation—mainly, ionizing forms. If a radiation-resistant organism similar to D. radiodurans were to be maintained as fully cryptobiotic in a desiccated habitat in near-surface martian environments, the SterLim team claims that the theoretical accumulated dose maximally allowed without overwhelming an ecological population might approach 20 kGy. At the surface of Mars, and 10 cm below the surface, the background dose rates for ionizing radiation range from 76-96 mGy/year;60 however, below several meters, the background radiation dose rate is significantly less. Therefore, the theoretical maximal survival dose of deeply frozen, desiccated D. radiodurans will be reached in Mars surface environments (i.e., 0-10 cm) in approximately 200,000 to 260,000 years. In contrast, ionizing radiation survival of deeply frozen, aqueous preparations of D. radiodurans is approximately four-times greater than when desiccated.61,62 It follows, in deeply frozen, aqueous conditions on the planet, that the theoretical ionizing radiation maximal dose (~80 kGy) in martian near-surface environments would be reached in approximately 1 million years (i.e., 80,000/0.076).
___________________
55 M. Ravikumar, R. Ravichandran, S. Sathiyan, S. Sudhakar Supe, 2004, Backscattered dose perturbation effects at metallic interfaces irradiated by high-energy x- and gamma ray therapeutic beams, Strahlentherapie Onkologie 180:173-178.
56 K. Kanjana, P. Ampornrat, and J. Channuie, 2017, Gamma-radiation-induced corrosion of aluminum alloy: low dose effect, Journal of Physics: Conference Series 860:012041.
57 V.M. Dekov, V. Arnaudov, F. Munnik, T.B. Boycheva, and S. Fiore, 2009, Native aluminum: Does it exist? American Mineralogist 94:1283-1286.
58 J.K. Fredrickson, S.W. Li, E.K. Gaidamakova, V.Y. Matrosova, M. Zhai, H.M. Sulloway, J.C. Scholten, M.G. Brown, D.L. Balkwill, and M.J. Daly, 2008, Protein oxidation: Key to bacterial desiccation resistance, ISME Journal 2:393-403.
59 Ibid.
60 See Table 3 in D.M. Hassler, C. Zeitlin, R.F. Wimmer-Schweingruber, B. Ehresmann, S. Rafkin, J.L. Eigenbrode, D.E. Brinza, et al., 2014, Mars’ surface radiation environment measured with the Mars Science Laboratory’s Curiosity Rover, Science 343(6169):1244797.
61 R.C. Richmond, R. Sridhar, and M.J. Daly, 1999, Physicochemical survival pattern for the radiophile Deinococcus radiodurans: A polyextremophile model for life on Mars, SPIE 3755:210-222.
62 L.R. Dartnell, S.J. Hunter, K.V. Lovell, A.J. Coates, and J.M. Ward, 2010, Low-temperature ionizing radiation resistance of Deinococcus radiodurans and Antarctic Dry Valley bacteria, Astrobiology 10:717-732.
For microorganisms directly on the surface of Mars or its moons, ultraviolet light and charged particles govern survival.63 However, just below the surface (>5 mm), extending several meters down, much more penetrating ionizing radiation predominates.64 For all cosmic radiation forms, the martian atmosphere attenuates dose rates by a factor of two.65 However, because Phobos and Deimos lack atmospheres, the theoretical maximal ionizing radiation dose would be reached twice as fast, in approximately 125,000 years when desiccated at their surfaces. This sterilization value can be accepted with a high degree of certainty, as there would be no possibility of a reconstituting event, such as hydration, to revive a microbial population on Phobos or Deimos. Moreover, the temperature of the upper few cm of Phobos and Deimos regolith intermittently reach or exceed 273 K, the freezing point of water (see Table 1.1), caused by solar heating. In vacuo, this diurnal temperature cycling causes desiccation, which is bactericidal to even the most radiation-resistant microbes in a matter of months at temperatures above the freezing point of water.66 It is a significant omission that neither the SterLim nor the JAXA teams accounted for the desiccating effect of diurnal temperature changes. Chapter 3 will return to this again because it will be an important factor in determining whether or not samples from the martian moons are designated restricted or unrestricted Earth return.
PHOBOS/DEIMOS SURFACE REFORMATION BY NATURAL METEOROID IMPACTS
The SterLim team noted that the turnover due to meteorite collision on the lunar surface (i.e., impact gardening) occurred with a confidence of 50 percent down to a depth of ~2 cm at least once every 10 million years. Thus, given that the meteorite flux at the martian system is even lower that at the Moon, the SterLim team concluded that the effect of impact gardening can be ignored.
The JAXA team showed that the collision of martian rocks forms a global thin layer (<0.1 mm) of materials contaminated with martian rock fragments on the surface of the moons, as previously mentioned. (See “Distribution of Mars Ejecta Fragments by Impacts, Recirculation, and Reimpact,” earlier in this chapter.) This layer is radiation (UVC and ionizing forms) sterilized in a short time (i.e., a few thousand years). However, if part of the layer is covered by an ejecta blanket due to a collision from interplanetary space, the blanket acts as a radiation shield. From the size-frequency distribution of martian craters,67 the size-frequency distribution of the interplanetary impactors to the Mars system was estimated using the pi-group, crater scaling law. Then the impact flux on the satellites was estimated using the cross-sectional area ratio of the satellites and Mars. The thickness of ejecta blanket as a function of the distance from the impact point was obtained based on the pi scaling. The fraction of the shielded area with the blanket of thickness 3 mm or more (the area for which radiation sterilization was not completed within 100,000 years) was found to be about 0.1 percent.68
Committee Assessment
The final factor in the chain of potential sterilizing process is the reformation of Phobos’s or Deimos’s surface by the natural fall of meteoroids from interplanetary space. However, this effect is insignificant due to the low flux of impactors.
In addition to bombardment by meteoroids, the surface of Phobos and Deimos can be physically weathered by thermal fatigue.69 This mechanism breaks down rocks into smaller pieces, thus exposing new surface area and
___________________
63 D. Matthiä, D.M. Hassler, W. de Wet, B. Ehresmann, A. Firan, J. Flores-McLaughlin, J. Guo, et al., 2017, The radiation environment on the surface of Mars: Summary of model calculations and comparison to RAD data, Life Sciences in Space Research 14:18-28.
64 D.M. Hassler, J.W. Norbury, and G. Reitz, 2017, Mars Science Laboratory radiation assessment detector (MSL/RAD) modeling workshop proceedings, Life Sciences in Space Research 14:1-2.
65 D.M. Hassler, C. Zeitlin, R.F. Wimmer-Schweingruber, B. Ehresmann, S. Rafkin, J.L. Eigenbrode, D.E. Brinza, et al., 2014, Mars’ surface radiation environment measured with the Mars Science Laboratory’s Curiosity Rover, Science 343(6169):1244797.
66 J.K. Fredrickson, S.W. Li, E.K. Gaidamakova, V.Y. Matrosova, M. Zhai, H.M. Sulloway, J.C. Scholten, M.G. Brown, D.L. Balkwill, and M.J. Daly, 2008, Protein oxidation: Key to bacterial desiccation resistance, ISME Journal 2:393-403.
67 See, for example, W.K. Hartmann, 2005, Martian cratering 8: Isochron refinement and the chronology of Mars, Icarus 174(2):294-320.
68 See Table 7.2 in Fujita et al., 2018, “Assessment of Microbial Contamination Probability for Sample Return from Martian Moons.”
69 M. Delbo, G. Libourel, J. Wilkerson, N. Murdoch, P. Michel, K.T. Ramesh, C. Ganino, C. Verati, and S. Marchi, 2014, Thermal fatigue as the origin of regolith on small asteroids, Nature 508:233-236.
making fresh regolith. This thermal fragmentation is predicted to occur at a rate orders of magnitude faster than fragmentation due to micrometeoroid impact. Although the effect was not considered by either the SterLim or the JAXA team, it could significantly enhance the rate at which any biological materials present are exposed and degraded by radiation.














