
6
Recovery of Marine Ecosystems
6.1 OVERVIEW
In recent years, there has been increased attention to nature- or ecosystem-based solutions in confronting climate change. In 2005, enhanced forest conservation (reducing emissions from deforestation and forest degradation) was promoted as a global initiative for greenhouse gas removal in developing countries (Angelsen et al., 2009). The United Nations (UN) started a Trillion Tree Campaign in 2007 to rehabilitate degraded land. In marine systems, much of the focus has been on coastal ecosystems, including mangroves, salt marshes, and seagrasses, perhaps because they can be managed in ways similar to those for terrestrial systems (NASEM, 2019). These coastal blue carbon systems largely comprise vascular plants, and several research efforts are already under way to study these systems and their potential for long-term C storage.
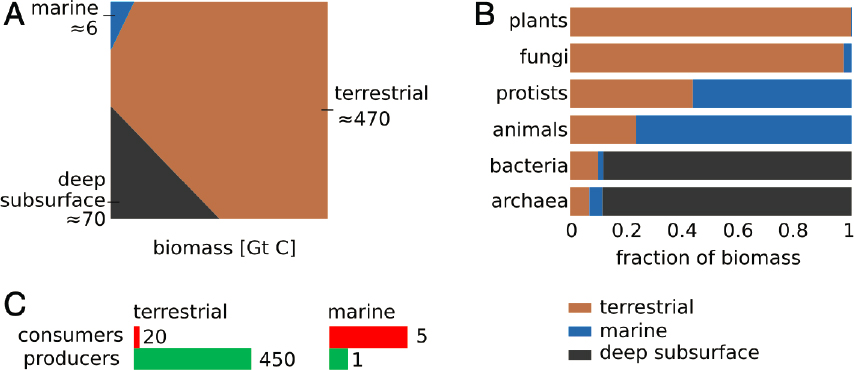
To date, much of the discussion around ecosystem-based solutions has focused on terrestrial and urban context, with risk reduction, such as the buffering ability of salt marshes and mangroves, on a local level (Solan et al., 2020). Benthic, pelagic, and offshore ocean systems are an emerging area of interest, yet there is a considerable amount of uncertainty about the potential for these marine systems to store and sequester carbon. Unlike terrestrial systems, a large fraction of the biomass in marine systems generally comprises animals, with consumers, including microbes and protists, far outweighing producers (Figure 6.1). Many of these organisms, especially the largest predators, are at risk from human activities such as fishing, shipping, pollution, introduction of invasive species, and habitat destruction. Although the fraction of biomass carbon in the oceans is relatively small compared to that of terrestrial systems, the role of animals in biogeochemical cycles and ecosystem structure has garnered attention, especially in light of the widespread impact of human activities on the state of the oceans and the co-benefits of ecosystem recovery approaches. At the same time, studies of kelp and other macroalgae such as Sargassum, benthic algae, and phytoplankton, have increased our understanding of their potential role in the carbon cycle.
The interconnected nature of climate change and biodiversity loss means that resolving either issue requires consideration of the other (Pörtner et al., 2021). In this chapter, we use the term “ecosystem-based” solutions to avoid concerns about the framing of the terms “natural,” “nature-based
solutions,” or NBS (Bellamy and Osaka, 2019). As in terrestrial systems, extensive monocultures of macroalgae, for example, could come at the expense of marine biodiversity. Yet there have been some important policy guidelines that have emerged from the NBS literature:
- Ecosystem-based solutions are not a replacement for the rapid phase-out of fossil fuels and must not postpone critical action to decarbonize our economies.
- They encompass the protection, restoration, and management of a wide array of natural and semi-natural ecosystems on land and in the sea.
- They are created, employed, managed, and monitored in collaboration with Indigenous peoples and local communities, respecting local rights and generating local benefits.
- Ecosystem-based solutions potentially support or enhance biodiversity from the level of the gene up to the ecosystem.1
Here, we focus on species and ecosystems in their native ranges, many of which have been depleted or altered. We do not discuss blue carbon efforts, which are often framed as coastal systems dominated by vascular plants and have been examined in earlier reports (NASEM, 2019). Efforts to enhance ecosystem functions, such as artificial upwellings and iron fertilization, are discussed elsewhere in this report and are not considered ecosystem-based in this context, though, of course, they both involve ecological responses to human interventions.
The protection and restoration of marine ecosystems and the recovery of fishes, whales, and other animals have the potential to aid in carbon dioxide (CO2) removal (CDR) and sequestration in the oceans, though considerable uncertainties remain, including the role of marine animals in nutrient cycles, the global areal extent of macroalgal and other ecosystems, and the fraction of carbon that is stored or sequestered during many of these ecological processes. See Section 1.4 for discussion of the ocean carbon cycle and the biological pump. In this chapter, we focus on manage-
___________________
ment actions that could result in ocean CDR, using a framework that compares natural, preindustrial processes to present-day fluxes, the difference being anthropogenic perturbation (see, e.g., Regnier et al., 2013). Rather than focus on present-day fluxes, the emphasis is on the restoration of natural processes and the impact of these activities on the carbon cycle.
Despite the uncertainty of these estimates, ecosystem-based solutions focused on conservation and recovery are likely to be an attractive part of CDR approaches, in part because they offer low-regret solutions, with many perceived co-benefits and the potential for global governability (Gattuso et al., 2021). The United Nations, for example, has declared 2021 to 2030 as the UN Decade on Ecosystem Restoration.2 Critical to this approach will be taking a whole-ecosystem perspective, going beyond individual populations or species and considering the changes and responses on various timescales that will be necessary to understand the role of CDR in restoration and conservation.
According to criteria described in Chapter 1, the committee’s assessment of the potential for ecosystem recovery, as a CDR approach, is discussed in Sections 6.2–6.5 and summarized in Section 6.6. The research needed to fill gaps in understanding of ecosystem recovery, as an approach to durably removing atmospheric CO2, is discussed and summarized in Section 6.7.
6.2 KNOWLEDGE BASE
The ocean is a sink for about 25 percent to 30 percent of the atmospheric CO2 emitted by human activities, approximately 2.5 to 2.6 petagrams (Pg) C/yr (Gruber et al., 2019; Watson et al., 2020). For this carbon to be sequestered at the scale of multiple decades, it must be transferred through food webs, enter sediments, or sink below the surface layer to the deep ocean, generally more than 1,000 meters (Passow and Carlson, 2012; Martin et al., 2021).
The biological carbon pump, which results from photosynthetically produced organic matter being transported from the surface layer to depth, is responsible for a large portion of the observed vertical gradient of natural carbon in the ocean (see Figure 1.8). The complex pathways of the biological carbon pump are driven by numerous biophysical and chemical interactions, including phytoplankton productivity, zooplankton grazing, marine vertebrate interactions, oceanic mixing and turbulence, advection, and the sinking of particles and aggregates (Ducklow et al., 2001; Nayak and Twardowski, 2020; Martin et al., 2021). Where possible, we have included estimates for C sequestration below the permanent pycnocline, or deep ocean, though in some cases estimates are only available to depths of 100 meters or so.
The preindustrial ocean carbon cycle is thought to have been in approximate steady state, with the biological uptake and downward export of surface CO2 largely balanced by the return of respired CO2 from the deep sea by the physical circulation. Despite the large role of marine systems in the carbon cycle, our understanding of changes or perturbation in ocean CO2 storage from past degradation is not well constrained, and our knowledge of the effects on ocean C storage by restoring and protecting marine organisms, ecological functions, and ecosystems is still emerging. For too long, the disciplines of population biology (and, as a subset, fisheries management), ecosystem ecology, and biological oceanography have proceeded on separate paths. A first step toward integrating these approaches would be to examine the ecological and biogeochemical baselines of the preindustrial ocean, including animals, macroalgae, phytoplankton, microbes, and their functions, while at the same time exploring how these baselines have shifted in the present ocean. Uniting these disciplines is even more complex in a changing ocean, with shifting circulation and residence times and the associated problems of using traditional metrics to infer impacts of marine biology on C storage (Koeve et al., 2020). One of the benefits of examining the CDR potential of
___________________
oceans is that it could spark greater collaboration between these fields and break down barriers to interdisciplinary studies.
The ocean’s ability to retain and remove carbon from the atmosphere is well established, and there is an extensive international effort to document the physicochemical uptake of anthropogenic carbon into the global ocean (e.g., Talley et al., 2016a; Gruber et al., 2019). In the Second State of the Carbon Cycle Report, Hayes et al. (2018) estimated that coastal waters of North America take up 0.16 Pg C/yr (Figure 6.2; Fennel et al., 2018). In Table 6.1, we list a few of the studies that we are aware of that have attempted to quantify the carbon stored in marine biomass, in deadfall carbon, or through ecological functions such as trophic cascades and nutrient subsidies. Many of these cases refer to present-day biological fluxes and are not directly comparable to estimates of net ocean uptake of anthropogenic carbon.
For the purposes of this report, we focus on the role of restoring marine organisms and ecosystems as a method of removing and sequestering anthropogenic CO2 in the oceans. For many of these systems, there have been few efforts to quantify the C impact of anthropogenic change over the past decades or centuries, though this is changing for fisheries and whales. Mariani et al. (2020), for example, note that fisheries have reduced the C sequestration potential of large fish, preventing the sequestration of about 22 Mt C through blue carbon extraction. This estimate does not include nutrient transfer and other potential indirect effects, which deserve further study. Restoration, of course, will not result in any net gains in C storage and sequestration if existing species, habitats, and ecological functions are not maintained. Conservation, in the form of protecting and preserving marine ecosystems, will also need to occur to make any gains in CDR.
The collective impact of anthropogenic changes, including rising CO2 levels, changes in nutrient inputs, and ecosystem alteration, on C processing and exchanges along ocean margins is complex and difficult to quantify (Regnier et al., 2013; Fennel et al., 2018). In this report, we largely focus on the potential role of conservation policy and recovery on the carbon cycle and biological pump, but clearly changes that occur because of climate change and other human impacts can have large effects on the flux of carbon in the oceans. Steffen et al. (2015) reported that the risk of transgressing planetary boundaries, such as biosphere integrity, could destabilize the current state of Earth systems. There are also potential trade-offs between fisheries and food security (e.g., Sustainable Development Goal 14) and efforts to promote additional CDR by restoring ocean ecosystems to something that might resemble a pristine state.
Although ecological processes have been well studied in terrestrial and coastal systems, the role of large marine animals and other organisms in the carbon cycle is still relatively new. One compelling question is whether animals and macroalgae have functional impacts that are disproportionate to their biomass, much as mat-forming mosses and lichens play important regulatory roles in the water cycle and inhibit microbial decomposition in some land ecosystems (Smith et al., 2015). Covering roughly 3 percent of Earth’s land, moss-dominated peatlands store nearly 33 percent of all global terrestrial carbon, about 540 Gt C (Turetsky, 2003; Yu et al., 2011).
Although marine ecosystems have been proposed as a climate solution, there is a fair amount of uncertainty surrounding our understanding of the ocean’s ability to store and sequester additional organic carbon, both in scale and in permanence. Given that ocean recovery is likely to be among the most publicly acceptable CDR approaches in the ocean, an approach with co-benefits and low regrets (Gattuso et al., 2021), it is likely to continue as a fruitful area of study. Yet how does it scale relative to other CDR schemes? Below we discuss a few of the mechanisms and systems that have been proposed as ecosystem-based solutions or areas that could merit further study.
TABLE 6.1 Selected Studies of Carbon Stored or Sequestered in Existing or Restored Ecosystems
| Mechanism | System | Region | C Stored or Sequestered | Reference |
|---|---|---|---|---|
| Macroalgae flux to detritus | Macroalgal–sediment systems | UK | 0.70 Tg/yr | Queirós et al., 2019 |
| Coralline algae growth and deposits | C storage by coralline algae and the beds they create | Global | 1.6 × Gt C/yr | van der Heijden and Kamenos, 2015 |
| Potential C sequestered by global macroalgae | Export of C to the deep sea and burial in coastal sediments | Global | 173 Tg C/yr | Krause-Jensen and Duarte, 2016 |
| Global ocean C export | Biological pump, export of biogenic particles from surface waters to stratified interior | Global | ~6 Pg C/yr | Siegel et al., 2014 |
| Diel vertical migration of fish and other metazoans | Global C fluxes and sequestration by fish and metazoans due to respiration, fecal pellets, and deadfalls | Global | 5.2 Pg C/yr | Pinti et al., 2021 |
| Gelatinous zooplankton C flows | Sinking of jelly-falls and fecal matter below 100 meters | Global | 1.6–5.2 Pg C/yr | Luo et al., 2020 |
| Fish-based contributions to ocean C flux | Passive and active flux from fish feces and migration | Global | 1.5 ± 1.2 Pg C/yr | Saba et al., 2021 |
| Recovery of marine vertebrates and increase of wild biomass | Role of vertebrates in the ocean C cycle, including living biomass, deep-sea carcasses, and nutrient cycling | Global | No total provided | Martin et al., 2021 |
| Baleen whale biomass and carcasses | Storage of C in living whales and sequestration of C in deep-sea whale carcasses | Global | 0.0089 Gt C/yr, with potential for significant increase in restored populations | Pershing et al., 2010 |
| Sea otter and kelp forest trophic cascade | Increase in C storage by kelp forests via sea otter suppression of herbivorous sea urchins | Northwest North America | 4.4–8.7 Tg C total stored | Wilmers et al., 2012 |
Macroalgae
Macroalgae form extensive and highly productive benthic marine habitats. Yet macroalgae afforestation has largely been overlooked as a natural source of sequestration in blue carbon schemes, in part because of challenges in documenting the amount of carbon that is sequestered during particulate organic carbon (POC) export (see Figure 5.1). The flowering plants that make up mangrove, seagrass, and salt marsh ecosystems have rhizomes and roots that can retain and store carbon in the soil for centuries. In contrast, macroalgae tend to be shorter lived, with all of their biomass growing above the seafloor: kelp typically have holdfasts attached to rocky surfaces, and planktonic seaweeds such as Sargassum typically float near the surface. Despite these differences, the carbon that macroalgal systems supply to sediment stocks in angiosperm habitats has been included in earlier blue carbon assessments, so they have already been recognized as playing a role in CDR (Krause-Jensen et al., 2018). In this section, we discuss the protection and restoration of native seaweeds. Seaweed cultivation and sequestration CDR is discussed in Chapter 5. Given that our understanding of seaweed aquaculture is related to natural macroalgal systems, see the knowledge base section of that chapter for further discussion of macroalgae and CDR.
Among the largest and most widely distributed macroalgae are the brown algae known as kelps and rockweeds (orders Laminariales, Tilopteridales, Desmarestiales, and Fucales), growing in dense populations or forests (Duffy et al., 2019). Although they cover about 28 percent of the world’s coastlines, kelp forests are declining faster than coral reefs and tropical forests (Feehan et al., 2021). Approximately 82 percent of kelp productivity becomes detritus (Krumhansl and Scheibling, 2012), and much of the carbon in macroalgae is assumed to return to its inorganic form in the water column through herbivory. Sinking speed of detritus depends on tissue type, size, and whether it has been consumed by herbivores (Wernberg and Filbee-Dexter, 2018). Export depths and sinking rates can vary by several orders of magnitude. Kelp forests in high-energy environments have a high potential for long-distance export, especially in areas with steep grades to depth (Wernberg and Filbee-Dexter, 2018).
In their study of the role of macroalgae in C sequestration, Krause-Jensen and Duarte (2016) estimated that about 173 Tg C/yr (or 11 percent of net primary production) could potentially be sequestered in marine sediments and deep-sea waters. They further suggest that roughly 70 percent of the sequestered carbon is due to the export of recalcitrant dissolved organic carbon (DOC) to the deep sea. The scale of this sequestration is comparable to all other coastal blue carbon habitats combined (Duarte et al., 2013). Note that Krause-Jensen and Duarte (2016) calculated the amount of carbon that could be sequestered if all ocean regions with enough light on the seafloor grew macrophytes. Hence, these estimates do not necessarily reflect the long-term sequestration by present-day macrophyte populations. Future research could refine these calculations for macroalgae afforestation sequestration in different oceans and ecoregions.
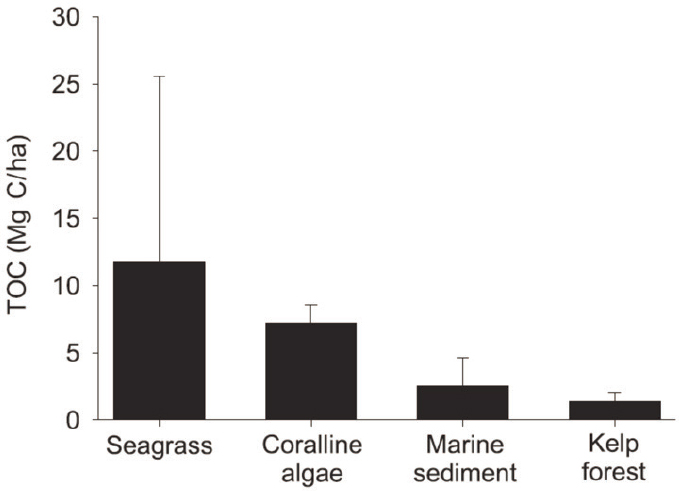
Queirós et al. (2019) estimated that the magnitude of detritus uptake within the deep-sea food web and sediments varies seasonally in the English Channel, with an average net sedimentary organic macroalgal C sequestration of 8.75 g C/m2, about 4–5 percent of estimates for mangroves, salt marshes, and seagrass beds. The average net sequestration per year of POC in sediments of the English Channel is 58.74 g C/m2, about 26–37 percent of blue carbon habitats (Figure 6.3). In a study of the Great Atlantic Sargassum Belt, Bach et al. (2021) found that biogeochemical feedbacks, nutrient reallocation, and calcification could greatly reduce the CDR efficacy of floating seaweed, though it could play an important role in increased ocean albedo.
Proposed restoration techniques include afforestation through seeding of macroalgae; the reversal of trophic cascades through the protection of sea otters and other predators; and exclusion methods such as flexible fencing to reduce sea urchin densities (Sharma et al., 2021).

Benthic Algae
Coralline or nonfleshy benthic algae have a worldwide distribution, comprising systems that can be thousands of years old (van der Heijden and Kamenos, 2015; Riosmena-Rodríguez et al., 2017). They have a large organic C stock (Figure 6.4) with the potential to store and sequester carbon. Yet relatively little is known about the role of calcified algae in C burial, including the balance between burial and the biological processes that release carbon (van der Heijden and Kamenos, 2015; Mao et al., 2020). Calcifying algal systems could represent a substantial, global-scale C repository with millennial longevity, though they are sensitive to climate variability and disturbance from trawling and other activities. The calcification process also reduces seawater alkalinity and increases the partial pressure of CO2, reducing the CDR efficacy of calcifying algae.
Animals and the Carbon Cycle
It has been suggested that conserving and restoring large marine vertebrates and other animals could provide an ecologically sound alternative to more intensive ocean CDR schemes such as iron
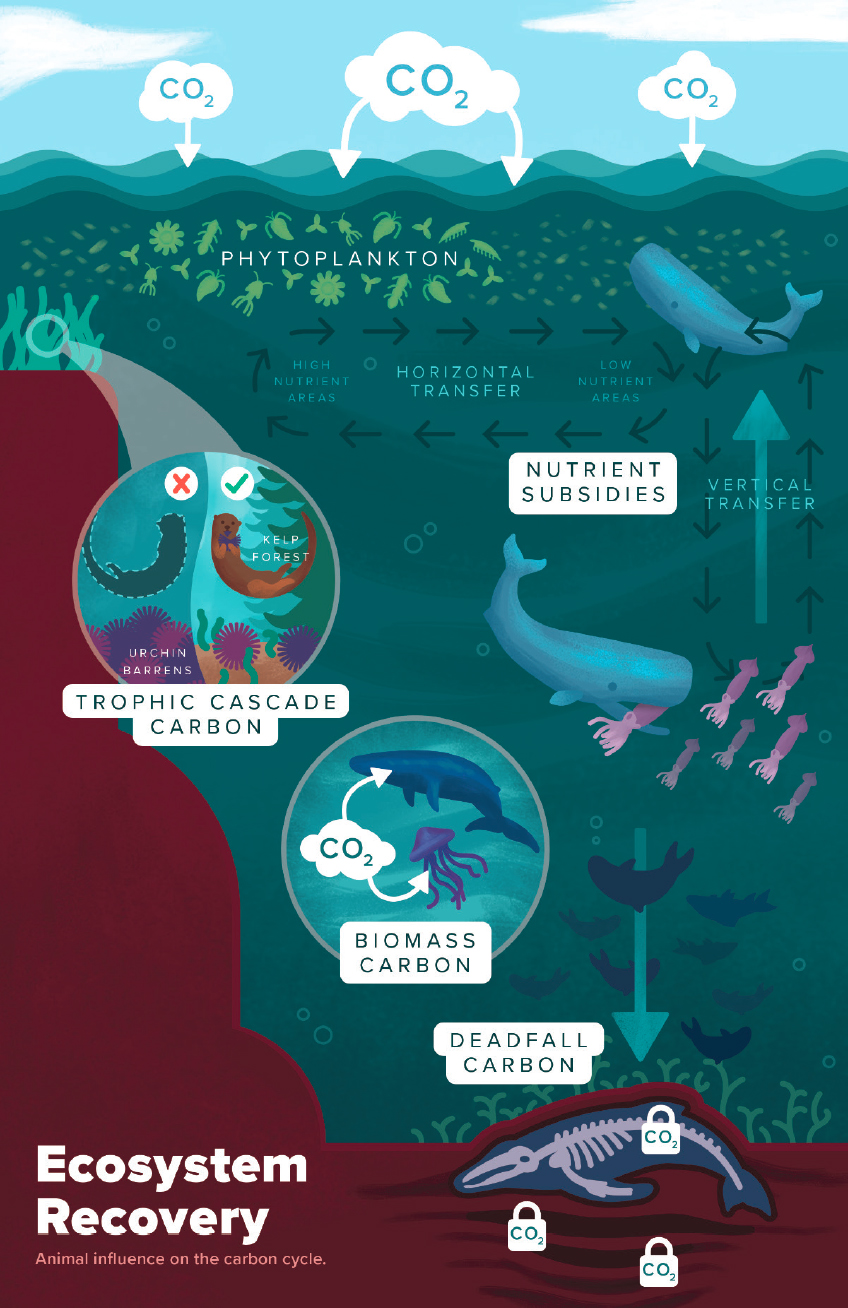
fertilization (e.g., Pershing et al., 2010). There are several ways that animals can influence the ocean carbon cycle (Figure 6.5): through the carbon stored in animal biomass; in carcasses and excretion, especially when they are exported to the deep sea; through nutrient subsidies and fertilization; and through trophic interactions. We briefly discuss these pathways in the sections below.
Animal Biomass
Carbon can be stored or sequestered in the living biomass of marine populations. Since marine animals can self-perpetuate through reproduction, they can continually generate new biomass. The carbon stored in recovering populations can be considered sequestered, as Martin et al. (2021) noted, “perhaps infinitely.” The rebuilding of fish stocks after decades of overfishing, and the increase of deadfall carbon, will reactivate a natural C pump driven by large marine fish (Mariani et al., 2020). Many of these fishing activities, especially on the high seas, would not be economically viable without subsidies. Bianchi et al. (2021) estimate that the pre-exploitation biomass was 3.3 ± 0.5 Gt, cycling roughly 2 percent of global primary production and producing 10 percent of surface biological export. The ecological impact of recovery can enhance or reduce the amount of carbon stored in ecosystems, depending on its ecological function and whether a species is an herbivore or predator (Schmitz et al., 2018).
Bar-On et al. (2018) estimated that marine animals comprise approximately 1.4 Gt C (Figure 6.1). This includes whales and other marine mammals, fishes, and other vertebrates; zooplankton; benthic species; and other invertebrates. (It does not, however, include echinoderms, one of the most diverse groups of marine animals, and is thus likely an underestimate.) Although we could not find total estimates for marine mammals, marine arthropods comprise about 1.0 Gt C, and fish (freshwater and marine) comprise about 0.7 Gt C. Pershing et al. (2010) estimated that rebuilding populations of eight whale species would store and sequester 8.7 Mt C in living biomass. Although these numbers are relatively small compared to terrestrial plant-based carbon, many animal populations in the ocean have been overexploited or reduced because of indirect human activities such as
pollution and habitat destruction. The recovery of historic numbers of marine animals presents an opportunity in terms of biomass, deadfall carbon (see below), and the restoration of zoogeochemical pathways in the carbon cycle (Schmitz et al., 2018).
There have been a few attempts to calculate these values for vertebrates, for example, an assessment of oceanic C for the United Arab Emirates for fishes, marine mammals, sea turtles, and seabirds (Pearson, 2019). But a global assessment of the biomass and geochemistry of marine animals in the present, past, and future under various recovery scenarios has not been completed.
Deadfall and Excreted Carbon
The primary movement of carbon from surface waters to the stratified interior is through the biological pump, which involves the sinking of biogenic particles that range from marine snow to metazoans, such as fish and zooplankton, and their feces (e.g., Falkowski et al., 1998). Active vertical migration also has a large impact on the C export flux of the biological pump, with organisms moving large distances between the surface and depths, potentially enhancing C export (Archibald et al., 2019). The carbon in carcasses and excreted material becomes available to food webs or bacteria.
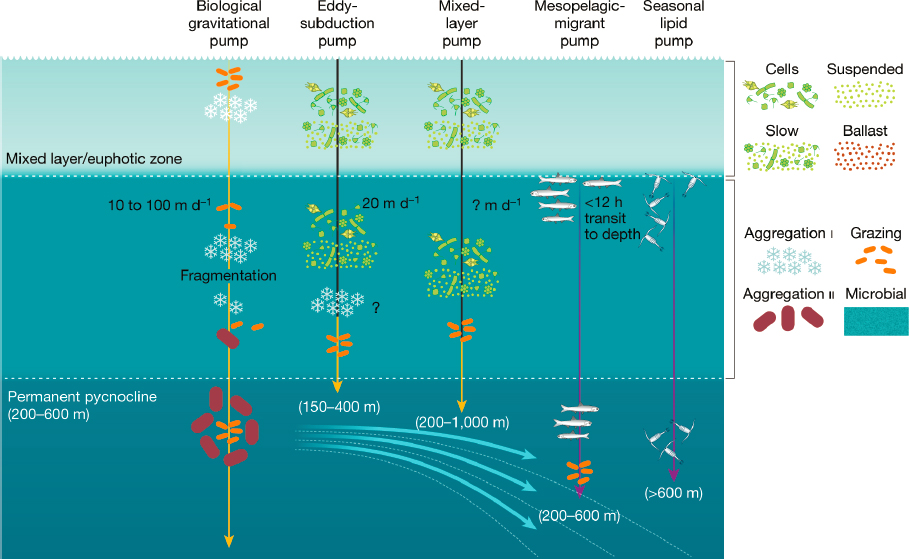
Boyd et al. (2019) estimated that the mesopelagic migrant pump could export more than 2 Pg C/yr. The concept for this pump is based on long-established observations of diurnal migration in zooplankton and fishes, the movement of the animals beneath the permanent pycnocline, and the release of fecal pellets at depth, with rapid sinking rates (Figure 6.6). In another study, Saba et al. (2021) calculated that fish contribute 16 percent ±13 percent to the global downward flux of carbon, equivalent to 1.5 ± 1.2 Gt C/yr. Whale falls, or cetacean carcasses that sink to the deep seafloor can sequester carbon for hundreds to thousands of years. Pershing et al. (2010) estimated that although relatively small in present numbers, especially when compared to the global C sink,
whale falls would remove 1.9 x 105 t C/yr if baleen whale stocks were restored. Recently, gelatinous zooplankton, or jellyfish such as cnidarians, ctenophores, and tunicates, have received some attention as potentially fast and efficient conduits of carbon-rich organic biomass into the deep ocean (Lebrato et al., 2019). Luo et al. (2020) estimated that the sinking of jelly-falls and fecal matter below 100 meters equals about 1.6–5.2 Pg C/yr, and 0.6 to 3.2 Pg C reaches 1,000 meters, the generally accepted depth for long-term sequestration.
Nutrient Subsidies and Fertilization
Ocean fertilization—adding nutrients (usually iron) to the upper ocean to stimulate plankton blooms and remove CO2—is perhaps the most well-known and highly debated method of ocean CDR (see Chapter 3). Whereas artificial iron fertilization has been relatively well studied, research on the role of marine animals in nutrient fluxes (natural fertilization) and its potential impact on C storage and sequestration is for the most part still emerging.
Relief of iron deficiency in areas of the sub-Antarctic Southern Ocean can enhance C export into the deep ocean and sediment (Pollard et al., 2009), and natural supplies of iron appear to result in much higher export efficiency than artificially added iron (see Chapter 3). Such findings on natural supplies of iron, along with recent research on the high iron content in the feces of whales, salps, and other marine animals, have prompted several efforts to quantify the role of marine animals in the iron cycle and other biogeochemical cycles, such as nitrogen and phosphorus (Lavery et al., 2010; Roman et al., 2014; Ratnarajah et al., 2018; Böckmann et al., 2021). For species that feed at the surface, such as many baleen whales, the recycling of iron and other nutrients likely aids in the retention of autochthonous, or recycled, iron, but does not necessarily support new production (e.g., Lavery et al., 2014).
Animals such as large fishes, sea turtles, marine mammals, and seabirds might be specifically essential for the biogeochemical cycle of several elements because they are widespread, highly mobile, and globally distributed (Wing et al., 2014; Tavares et al., 2019). Sperm whales, for example, feed on deep-living prey and defecate at the surface, transporting allochthonous iron into the euphotic zone and raising the nutrient standing stock of surface waters (Lavery et al., 2010). Lavery et al. assumed that about 75 percent of the total iron defecated by sperm whales persists in the photic zone, with iron in the beaks of cephalopod prey sinking. This nutrient contribution can enhance new production, increasing net uptake of CO2 from the atmosphere and C export to the deep ocean, though the amount of carbon that is stored, sequestered, or released back into the surface waters remains an area of active debate (for a review of marine vertebrates in the carbon cycle, see Martin et al., 2021; Roman et al., 2021).
Many baleen whales are capital breeders, storing resources in highly productive high-latitude areas, for annual migration to lower-latitude and often oligotrophic systems where they breed. They can transport nutrients between these systems via urine, placentas, carcasses, and sloughed skin (Roman et al., 2014). These can be considered external, or allochthonous, nutrient inputs that could ultimately lead to new production and C sequestration, because nutrients are released at locations thousands of kilometers from their origin, often in areas that are nutrient limited. (Areas that are not nutrient limited may not have a large impact on the global total.) Nitrogen released by the modern-day population of Southern Ocean blue whales on their calving grounds, for example, is estimated to result in the storage and sequestration of 5.1 x 102 t C/yr; yet before whaling greatly reduced this species, estimates would have been closer to 1.4 x 105 t C/yr (Table 6.2; Roman et al., 2014).
Despite the attention given to marine mammals, it is likely that the far more abundant krill, salps, and other zooplankton play an important role in the biogeochemical cycles. Vertical movement by marine organisms has the potential to increase ocean mixing across pycnoclines, though these contributions might be small when compared to internal gravity waves (Kunze, 2019). A
recent study found that fecal pellet material from salps released more bioavailable iron to Southern Ocean phytoplankton than krill pellets (Böckmann et al., 2021). One estimate suggests that about two-thirds of these salp pellets stay near the surface (Iversen et al., 2017). Phytoplankton communities took up to five times more iron from the salps’ fecal pellets than from krill feces. The variation in iron availability in different types of fecal pellets could influence biogeochemical cycles and C fixation of the Southern Ocean, potentially representing a negative feedback to climate change as salp populations increase with reduced sea-ice coverage (Böckmann et al., 2021). Such regime shifts are linked to changes in sea-ice coverage and are not considered part of an ecosystem recovery approach to CDR.
Trophic Interactions
Several studies have examined the food web dynamics of animals in the carbon cycle. Planktivorous fish and invertebrates can convert carbon into longer-lived biomass and rapidly sinking fecal pellets, increasing C export (Martin et al., 2021). At the same time, grazing animals such as dugongs and green turtles can decrease seagrass meadow structure and biomass, decreasing the quantity fated to the detrital pool (Scott et al., 2021).
Predators also play a role in these systems. A few studies have examined the role of predators in enhancing C sequestration, including sharks (predation risk) and sea otters (consumptive trophic cascade). The risk of predation by sharks limits herbivore abundance to zones close to reefs, resulting in reduced grazing, increased algal biomass, and improved sedimentary C stocks and sequestration away from reef refuges (Atwood et al., 2018). In a classic example, sea otters are considered keystone predators in the North Pacific, consuming sea urchins and other benthic species, which can have a positive impact on kelp and C storage and sequestration. Wilmers et al. (2012) estimated that sea otter predation could increase the storage of 0.015–0.043 Gt C/yr and the sequestration of 0.0013–0.023 Gt C/yr by restoring kelp forests. Gregr et al. (2020) predicted a net benefit of CAD$2.2M for the C sequestered by sea otters.
In vegetated coastal systems, Atwood et al. (2015) noted that there was sufficient evidence to suggest that intact predator populations can be vital to preserving or increasing carbon reserves stored in coastal or marine ecosystems. As such, policy and management need to be improved to reflect these realities. Beyond these coastal systems, there is a need for a deeper exploration of the role in trophic interactions on macroalgae as well as in pelagic and offshore benthic ecosystems.
Ecosystem-Based Solutions
Ecosystem-based solutions, also known as nature-based solutions, have the potential to protect natural habitats and restore modified ecosystems while providing other societal benefits for humans, such as health and ecosystem services, and protecting other species (Solan et al., 2020). Ecosystem-based solutions to climate change rely largely on traditional conservation efforts to protect and restore marine ecosystems and native species. Roman et al. (2015), Duarte et al. (2020), and others have recently reported on the ongoing recovery of many large marine animals, including humpback whales, sea turtles, and elephant seals, after decades of overharvesting. A variety of measures, including reduction of direct and indirect take, expansion of protected areas, and improved enforcement have helped many of these species, and dedicated efforts have the potential to restore the diversity of marine life.
Although the overriding theme of this chapter is CDR in the ocean, it would be counterproductive to ignore the role of marine conservation efforts to protect current stores and flows in the carbon cycle. Lovelock et al. (2018) noted that two steps are necessary for C sequestration in coastal systems such as mangroves, a process that likely holds true for pelagic and benthic systems: con-
servation is necessary to avoid additional CO2 emissions from further habitat loss, and restoration is used to restore C pools and, during their buildup, act as CO2 sinks. Similar steps are in play in protecting benthic and pelagic systems: to avoid leakage, current C removal processes must be protected, while others are restored. Benthic algae and sediments are at risk from bottom trawling, for example. Leaving sediments undisturbed from bottom trawling can protect stored carbon and enhance anoxic remineralization, which in turn generates alkalinity and reduces partial pressure of CO2 (pCO2). At the same time, new algal systems can be restored.
One of the big questions for any ecosystem-based research agenda is how much additional carbon is expected to be removed as a result of habitat restoration and population recovery. Answering this will require tests of concepts for the CDR potential of restoring benthic and pelagic systems. Oreska et al. (2020) showed that restoring seagrass meadows in Virginia removed 9,600 tons of CO2 from the atmosphere over 15 years, but it also heightened methane (CH4) and nitrous oxide (N2O) production, releasing 950 tons of CO2 equivalents. The meadow now offsets 0.42 t CO2 equivalents/ha per year. Similar efforts to examine the removal and release of CO2 and other greenhouse gases in different systems will be necessary to estimate the sequestration rates of restored marine ecosystems. In some cases, new methods for greenhouse gas accounting will likely be needed to calculate the net benefit of marine systems that include macroalgae, phytoplankton, metazoans, and microbes.
Marine Protected Areas and Habitat Restoration
The protection of marine ecosystems has typically followed two paths: (1) management of individual species or groups of species, such as the Marine Mammal Protection Act in the United States and protections under the International Union for Conservation of Nature (IUCN) and the International Whaling Commission, and (2) protection of habitats and communities through marine protected and conservation areas and other types of restrictions on habitat use. Developing effective marine protected areas (MPAs) for biodiversity and CDR requires identifying vulnerable habitats and restoring areas that will be the most effective inside and outside of protected areas. There is a rich literature on the value of spillover effects, networks, and dispersion capability for MPAs, but the focus on carbon is relatively new.
Roberts et al. (2017) discussed the ability of protected areas to help ecosystems and people adapt to the impacts of climate change by helping to mitigate sea-level rise, intensification of storms, shifts in species distribution, and other expected changes. Protected areas can also play direct and indirect roles in the many processes discussed in this chapter: promoting the storage of carbon in biomass and carcasses, protecting the seafloor from trawling and mining, protecting and restoring food webs and reversing human-induced trophic cascades, and reducing the pressures of overharvesting.
The push to protect more of the oceans was adopted by the IUCN World Conservation Congress in 2016, calling for 30 percent protection by 2030. This effort and similar ones proposed by the Biden administration in the United States and elsewhere provide an opportunity for researchers to examine the role of benthic and pelagic species in CDR. As populations and ecosystems change as a result of protections, expected increases in C sequestration can be measured using established and emerging technologies. To be effective, and to understand the role of protected areas in CDR, MPAs will need resources for monitoring and enforcement (see Monitoring and Verification below).
In addition to MPAs and habitat restoration, restrictions on destructive benthic and deep-sea activities may be necessary to retain carbon already captured in the seafloor. Marine sediments store approximately 2,322 Pg C in the top 1 meter, nearly twice that of terrestrial soils (Atwood et al., 2020). Seventy-nine percent of the global marine C stock is in abyss and basin zones that have largely been undisturbed. Areas that have been extensively modified have likely released a fair amount of carbon already. Only about 2 percent of C stocks is located in fully protected areas,
where it is safe from legal disturbance of the seafloor (Atwood et al., 2020). Recognizing the need to protect deep-ocean ecosystems and biodiversity, the IUCN recently called for a moratorium on seabed mining.3 The restriction or prohibition of seabed mining and bottom trawling will likely help store and retain old carbon and sequester new carbon in the seafloor.
Macroalgae Protection and Restoration
Although there are no global estimates for loss of macroalgal habitats, one study showed declines in kelp forests in 38 percent of ecoregions, with other areas showing increases, indicating that there is high geographic variability (Krumhansl et al., 2016). Pessarrodona et al. (2021) showed a loss of forest-forming seaweeds and rise of turf algae across four continents, resulting in the miniaturization of underwater habitat structure. Protection and restoration can avoid global losses in many of these ecoregions. Some losses of kelp, however, can be attributed to increasing water temperatures. In such cases, restoration and protection might not help retain kelp forests, and other methods such as assisted migration of kelp at the leading edge of their range might be necessary to increase kelp forests. Several other algae species, such as the commercially important Gelidium, a red seaweed that is the source of laboratory agar, have been overharvested and are subject to trade restrictions and regulations (Callaway, 2015).
There are several approaches to restoring degraded seaweed habitats and protecting existing ones, with a focus on CDR. Each of the three items below focuses on CDR, but they all are expected to enhance biodiversity and some ecosystem services, such as tourism and finfish fisheries.
Habitat Protection
Protecting areas with extant macroalgae and restoring historic areas is the first step to enhancing algal growth and CDR. Measures can include MPAs, reduced nutrient loading, reduced disturbance of the seafloor, and bans on extraction of stones and rocks from sensitive areas.
Protection of Carbon Sinks in Sediments Beyond the Habitats
Carbon export and sequestration from macroalgae typically depends on POC export to the seafloor. Trawling and seafloor mining can release this carbon back to the surface, so protecting these areas from human disturbance is essential in the CDR process.
Restoration of Macroalgae
In addition to protection, restoration of disturbed macroalgal systems is essential to the CDR approach. Methods can include the control of herbivores, such as sea urchins (see Reversing Trophic Cascades and Restoring Food Webs below) and the seeding of macroalgae, including techniques such as “green gravel,” small rocks seeded with kelp (Fredriksen et al., 2020).
Japan probably has the most experience in kelp restoration (Figure 6.7; Duarte et al., 2020). In the United States, kelp restoration is typically linked to the return of predators, such as sea otters (Wilmers et al., 2012). The U.S. seaweed and kelp industry is small compared to production in East Asia, but there is rising interest in the market for human consumption (Janasie and Nichols, 2018). A new partnership in British Columbia among the Haida Nation, Parks Canada, and Fisheries and Oceans Canada, academic institutions, nongovernmental organizations (NGOs), and commercial
___________________
fishers is focused on two coastal restoration areas for recovery of kelp and sea otters (Goldman, 2019).

Fish and Fisheries Management
The discussion of food systems in relation to CO2 emissions has typically focused on terrestrial agriculture and land-use changes (e.g., Crippa et al., 2021). In contrast, fisheries management has generally looked at the expected response of fish stocks to global change and related uncertainties (e.g., Howard et al., 2013). The potential global fisheries catches, for example, are expected to decrease by about 3 million tons for each increase per degree Celsius of global surface warming, with a high number of species turnover (Cheung et al., 2016).
This discussion has recently begun to change. The majority of commercially targeted fish populations are predatory, and the removal of biomass and alteration of the ecological structure and function of the ocean, even under regimes that are considered sustainable such as maximum sustainable yield, are likely to have impacts on the ocean carbon cycle and storage (Yodzis, 2001; Spiers et al., 2016; Stafford, 2019). As such, strategies intended to increase harvest, by keeping populations at 50 percent of carrying capacity or lower, will likely be ineffective as a C strategy. In addition to the direct take from fisheries and habitat destruction, indirect take can affect whales and other marine mammals. Currently, about 11 percent of commercially captured fish is discarded (Gilman et al., 2020), and marine mammal bycatch endangers and limits the recovery of many populations (Read et al., 2006).
Several new studies have estimated the potential role of rebuilding fish populations in the ocean carbon cycle. Marine fisheries have released at least 0.2 Gt of carbon into the atmosphere since 1950, with greater than 43 percent of the carbon released by high seas fisheries coming from areas that would be economically unprofitable without subsidies (Mariani et al., 2020). These calculations include the amount of carbon that was extracted rather than sequestered by dead fish sinking to the deep sea (21.8 Mt C), the amount of carbon extracted from the ocean as living biomass (37.5 Mt C), and the carbon emitted from burning fossil fuels and processing fish on land (202.8 Mt C). Bianchi et al. (2021) estimated that fish cycled roughly 2 percent of global primary production (9.4 ± 1.6 Gt/yr) and produced 10 percent of surface biological export before industrial fishing. Sala et al. (2021) estimated that bottom trawling and dredging release 0.4 Pg C/yr. This rather large number needs to be reconciled with ocean inorganic CO2 budget estimates from Gruber et al. (2019) and other researchers. By expanding the value of fish to include greenhouse gas mitigation and CDR,
in the form of fishery industrial emissions, biomass, deadfall carbon, and trophic cascade carbon, it is possible that there will be a shift in values that can help restore wild fish populations, while also maintaining protein supplied by seafood.
Restoring Populations of Large Marine Organisms
Given their potential to store and sequester carbon, conserving large species, and the largest individuals within a species, should be a top priority (Pershing et al., 2010). Historical records, genetic studies, and food web models all suggest that apex predators, large fishes, and marine mammals were more abundant in the past (Ferretti et al., 2010; Magera et al., 2013; Christensen et al., 2014). Unlike terrestrial systems, marine ecosystems should be top-heavy because of complex food webs and top-down effects, with more biomass stored in large predators than in lower trophic levels; the current bottom-heavy trophic structure could be a result of widespread human defaunation of the ocean (Woodson et al., 2020).
Marine organisms, including predatory fish and marine mammals, are especially vulnerable to human activities in the ocean. In addition to direct threats from commercial fishing, marine mammals and other vertebrates are also at risk from ship strikes, ocean noise, plastics, and other types of marine pollution (e.g., Rockwood et al., 2017; López Martínez et al., 2021). The reduction of these risks provides an opportunity for restoration and experimentation. The restoration of large marine organisms, through fisheries management and mitigation, MPAs, pollution reduction, and other protective policies, can result in increased biomass, deadfall carbon, and nutrient transfer and subsidies.
Reversing Trophic Cascades and Restoring Food Webs
Animals perform a complex set of trophic and nontrophic interactions that can cascade through food webs and affect carbon processes (e.g., Schmitz et al., 2018). The conservation and recovery of marine keystone species have been proposed as one way of restoring and enhancing C sequestration in kelp forests and other habitats. The repair of trophic interactions between sea otters and sea urchins through otter recovery, for example, could lead to a 1,200 percent increase in kelp biomass carbon within the otter’s range in the North Pacific (Wilmers et al., 2012). The loss of predators such as sharks can have a large effect on C burial in coastal ecosystems, potentially reducing burial by up to 90 percent in some habitats (Atwood et al., 2015). The C stock of macroalgae growing in high-risk areas for herbivorous fish that are away from coral reefs is 24 percent higher than in areas close to reef refuges (Atwood et al., 2018).
The restoration of intact food webs and the reversal of human-induced trophic cascades can enhance or reduce C storage (Estes et al., 2011; Schmitz et al., 2018). The recovery of herbivorous species, while good for biodiversity conservation, might not increase carbon sequestration. The return of the green turtle to the Lakshadweep Archipelago in the Indian Ocean, for example, resulted in overgrazing of seagrass, reduced in seagrass fish diversity, biomass, and abundance, and major declines in sediment-stored carbon (Gangal et al., 2021).
6.3 EFFICACY
The protection of marine ecosystems—including protection, relocation, restoration, and the reduction of pollution and overexploitation—is feasible and durable (Gattuso et al., 2018). Yet there have been no comprehensive attempts to estimate the CDR potential of restoring entire marine ecosystems and only a few efforts to quantify the global impact of restoring and protecting taxa or functional groups: for example, for macroalgae (173 Tg C/yr; Krause-Jensen and Duarte, 2016) or
restricting certain destructive practices such as bottom trawling (0.4 Pg C/yr; Sala et al., 2021). In contrast to terrestrial and coastal systems, with a large literature on natural climate solutions, there has been little quantitative research on the impact of management tools, such as protected areas and harvest restrictions, on C fluxes in marine ecosystems.
Perhaps the most important step toward including ecosystem restoration in a CDR scheme is measuring the net greenhouse gas benefits for a given project. This would likely involve measuring the carbon that is stored in biomass, sequestered as deadfall or fecal carbon, or enhanced and sequestered via processes such as nutrient fertilization. The production of CH4 or N2O, or the release of CO2, through respiration should also be accounted for.
Additionality
Restoring marine ecosystems offers two clear and overlapping benefits: reducing biodiversity loss and restoring the role of marine organisms in the carbon cycle. Reduced harvest, restored habitat, and larger no-take zones will almost certainly sequester more atmospheric carbon, though the scale of many of these reductions remains debated and is an important area for new research. There trade-offs between commercial fishing, food security, and CDR will need to be considered in management plans. One challenge in quantifying additionality in the protection of marine fauna is that changes in protection in one area can result in increased fishing in others, and many fish and marine mammals are migratory, with their ranges shifting as temperatures change and prey populations move (Cheung et al., 2016). The monitoring of many migratory species will need to go beyond political boundaries.
Durability
One of the advantages of ecosystem restoration is that marine organisms can self-perpetuate through reproduction, generating new biomass: carbon can be stored in stable populations “perhaps infinitely” (Martin et al., 2021). As biomass continues to stay relatively stable, more carbon would be stored through death and deposition, resulting in a long-term C drawdown. This process would happen naturally if humans were not continually altering biogeochemical cycles. Unfortunately, these cycles are expected to change as a result of rising temperatures, ocean acidification, and ocean deoxygenation, with the C capture and storage potential of the oceans, along with productivity and biomass, likely to decline (Gruber, 2011). Changes in surface nutrients and phytoplankton and zooplankton biomass could also reduce total biomass and body size of fish and other vertebrates (Boyce et al., 2010; Britten et al., 2016).
One of the greatest risks to the CDR potential of marine ecosystems, and an existential risk to humans and many other animals, is the current extinction crisis combined with an increased atmospheric concentration of CO2. The fossil record shows major responses in the oceans during global C perturbations and mass extinctions. In benthic systems, responses include reduced diversity, decreased burrow size and bioturbation intensity, and extinction of trophic groups (Bianchi et al., 2021). Models developed by Tulloch et al. (2019) show predicted declines and local extinctions of baleen whales and krill in the Southern Ocean by 2100 as a result of climate change. In the case of widespread ecosystem shifts, there is a potential for rapid changes that could result in the reversal of CDR gains.
There is some evidence, however, that diverse and abundant fish communities are more resilient in the face of warming waters and other climate shifts (Sumaila and Tai, 2020). MPAs could help build resilience to climate change for coral reefs and other ecosystems (e.g., Chung et al., 2019). In addition to the effects of increased greenhouse gases, management and protection regimes can be subject to changes in policy, which would affect the permanence of C removal.
TABLE 6.2 Estimated Amount of Carbon Fixed, Stored, or Sequestered in Pre-exploitation and Modern Cetacean Populations
| Mechanism | Species | Region | Pre-exploitation C Estimate (N) | Modern C Estimate (N, year) | C Fixed, Stored, or Sequestered | Reference |
|---|---|---|---|---|---|---|
| Living biomass carbon | Pantropical spotted dolphin (Stenella attenuata) | Eastern Tropical Pacific | 5.9 × 104 t C (3.6 x 106) | 1.4 × 104 t C (8.6 × 105, 2006) | Stored | Martin et al., 2016 |
| Living biomass carbon | Spinner dolphin (Stenella longirostris) | Eastern Tropical Pacific | 2.4 × 104 t C (1.8 × 106) | 1.4 × 104 t C (1.1 × 106, 2006) | Stored | Martin et al., 2016 |
| Living biomass carbon | 8 baleen whale taxaa | Global | 2.0 × 107 t C (2.6 × 106) | 3.1 × 106 t C (8.8 × 105, 2001) | Stored | Pershing et al., 2010 |
| Deadfall carbon | 8 baleen whale taxaa | Global | 1.9 × 105 t C/yr (2.6 × 106) | 2.9 × 104 t C/yr (8.8 × 105, 2001) | Sequestered | Pershing et al., 2010 |
| Whale pump | Sperm whale (Physeter macrocephalus) | Southern Ocean | 2.4 × 106 t C/yr (1.2 × 104) | 4 × 105 t C/yr (1.2 × 103, 2001) | Fixed | Lavery et al., 2010 |
| Whale pump | Blue whale (Balaenoptera musculus) | Southern Ocean | 1.3 × 108 t C/yr (2.4 × 105) | 2.8 × 106 t C/yr (5.2 × 103, 2012) | Stored | Lavery et al., 2014 |
| Great whale conveyor belt | Blue whale | Southern Ocean | 1.4 × 105 t C/yr (3.4 × 105) | 5.1 × 102 t C/yr (4.7 × 103, 2001) | Stored and sequestered | Roman et al., 2014 |
NOTES: Mechanisms include storage in living biomass, sequestration in whale carcasses sinking to the deep sea, and nutrient transport (vertical pump and horizontal conveyor belt) via excretion and defecation of nitrogen, phosphorus, and iron. C values are gross and do not account for the amount of carbon respired by cetaceans. Exploitation refers to bycatch for dolphins and industrial whaling for all other cetaceans.
a Balaenoptera acutorostrata, B. bonaerensis, B. borealis, B. brydei, B. musculus, B. physalus, Balaena mysticetus, Eschrichtius robustus, Eubalaena spp., Megaptera novaeangliae. SOURCE: Courtesy of Heidi Pearson, University of Alaska Southeast, and Roman et al., 2021.
Monitoring and Verification
Monitoring and verification are essential to protection efforts, especially when they have particular goals such as CDR and biodiversity protection. One concern about CDR is that an extensive monitoring and evaluation system would require large infrastructure for a relatively small return. Should there be a minimum requirement to invest in monitoring and evaluation? The current ocean uptake of anthropogenic carbon from physicochemical process is approximately 2.5 Pg C/yr; if the CDR approaches we discuss can increase uptake on a petagram scale, monitoring programs such as the Global Ocean Hydrographic Investigations Program, or GO-SHIP, should be able to detect these changes.
All of the processes we discuss will require documenting that CDR is occurring. This process will require going beyond indirect calculation to demonstrate enhanced sequestration at sink sites (Krause-Jensen et al., 2018). On land, the Verified Carbon Standard is the most commonly used standard. It includes several requirements: the emissions reduction or removal must be real, measurable, permanent, unique, and additional. Whether these measures, designed for terrestrial systems, will apply in marine C schemes remains to be seen (Krause-Jensen et al., 2018). Mesoscale nitrogen and iron enrichment experiments could test if marine mammal and seabird feces effectively modify phytoplankton processes, enhancing diatom biomass and increasing atmospheric CO2 drawdown (Boyd et al., 2000; Pollard et al., 2009).
Environmental Monitoring of Marine Protected Areas
Solutions for Monitoring MPA Ecosystem Health
Good monitoring and data collection are essential if MPAs are to deliver their intended biodiversity and CDR outcomes. Monitoring ecosystem health and environmental stressors in offshore and remote MPAs is particularly challenging because of limited access for data collection. Custom solutions can be developed to fit individual geographic settings, environmental conditions, and conservation priorities. Satellite remote sensing approaches, for example, can be used to monitor the evolution and environmental conditions of new MPAs (Kachelreiss et al., 2014).
Offshore and Remote MPAs
Deep-ocean and seamount MPAs In the Northeast Pacific off Canada, there are deep-ocean cabled sensor networks that have provided real-time access to the Endeavour Hydrothermal Vents, which was Canada’s first MPA, established in 2009. Data products from this site were developed to support management of the Endeavour MPA (Juniper et al., 2019). Monitoring was geographically extended in 2018 to support surveys of offshore Pacific seamount MPAs using remotely operated vehicles, autonomous sensor platforms, multibeam mapping, and vessel traffic data products (virtual automatic identification system, or AIS, fences). Although cable-based sensor networks are limited in their geographic extent, existing systems should be used to deliver monitoring data products. All of the remote technologies and data products could be made available for long-term, site-specific ecosystem health monitoring of deep-ocean and seamount MPAs.
Regional-scale monitoring Management of other deep-ocean ecosystems requires monitoring of regional-scale threats, such as marine heat waves and ocean deoxygenation, in addition to site-specific monitoring. Regionally deployed flotillas of new-generation Argo profiling floats to monitor ocean temperature, dissolved oxygen, and other seawater properties could be used for delivering management data products. Ships of opportunity could be coordinated and engaged to ensure that
the floats remain in the regions of interest (e.g., Smith et al., 2019a). The ships themselves could also provide data and data products. These autonomous-sensor platforms profile between the sea surface and ocean depths every few days, measuring water properties and relaying data via satellite. Data and data products from decades-long “Line” surveys in regions (e.g., Line P in the Northeast Pacific) provide a historic framework for understanding long-term ocean change in areas where these surveys take place.
Marine mammal activity in remote protected areas Cabled and autonomous mooring networks can deliver marine mammal monitoring solutions, using underwater hydrophone systems that record whale sounds and ship noise, and AIS data that track vessel movements with a focus on data products that inform long-term management by reporting on monthly and annual levels of marine mammal and large vessel activity in MPAs. In the case of real-time operational use, these data can be used for compliance and enforcement. In areas of known marine mammal activity, virtual AIS fences could be implemented to identify ship traffic and inform ships’ captains to reduce vessel strikes.
Ecosystem Health Solutions: Coastal Zone MPA Monitoring
Coastal zone MPA monitoring could be delivered in the United States through the existing regionally based Ocean Observing Systems (IOOS Strategic Plan 2018–2022; IOOS, 2018). The Strategic Plan already includes delivery of the benefit to “more effectively protect and restore healthy coastal ecosystems,” and this could be expanded to include observations and measurements of ocean CDR efficacy and environmental impacts. In areas not covered by IOOS, community-based monitoring programs could be implemented. These approaches are especially important in Indigenous communities (see, e.g., Kaiser et al., 2019).
Ecosystem Health Solutions: MPA Data Products
State of the ocean data products The collection of long time-series data by offshore and coastal observing systems provides critical information for evaluating local ocean health trends and their relationship to regional ocean change. Some permanent observatory installations host core sensors for monitoring seawater properties and noise levels. These properties and their relationships to ecosystem health include
- temperature—informs on long-term warming trends and marine heat waves;
- salinity (salt concentration)—detects anomalies in freshwater input to coastal waters;
- pressure (depth)—informs on sea-level rise, tsunami and storm surge events, tidal cycles; and
- dissolved oxygen concentration—key to detecting hypoxia events and long-term oxygen depletion that can have deleterious impacts on marine ecosystems.
Daily noise index data product Noise generated by surface vessels is well known to be harmful to marine mammals, and there is growing evidence of negative impacts on fish and invertebrates. To capture ship-source signature noise profiles, multihydrophone arrays are needed to ensure that the data meet ANSI standards. Once source signatures are captured, these data can be combined in real time to model the noise in heavily trafficked areas (Zhang et al., 2020) and potentially could be used to reduce noise through voluntary or mandatory measures. Long-term fixed hydrophone networks can provide a benchmark to assess increases or decreases in noise (Thomson and Barclay, 2020).
Biodiversity and stock assessments Information on trends in species diversity and fish abundances is often a key metric for monitoring conservation objectives in MPAs and plays an important role in understanding the carbon dynamics of these systems. Imagery from fixed observatory underwater cameras and regular ship-based video transect surveys could be used to develop data products on species diversity and abundance to inform MPA management and CDR research. Manual, machine vision, and crowdsourcing tools are several approaches that extract biological information from video and photo archives to reveal trends in species numbers (Matabos et al., 2017).
Solutions for Compliance and Enforcement Monitoring
Tools that deliver summary and real-time data products of vessel and marine mammal activity in MPAs, using data from vessel AIS and passive acoustic monitoring (hydrophone) data from cabled and autonomous listening stations, are important tools to support monitoring for compliance and enforcement. Hydrophone systems are expensive to invoke in all regions, but research is under way to use satellite data for detection of some marine mammals that could fill large areas (Platonov et al., 2013). Summary data products will permit monthly review of vessel and marine mammal activity in remote and coastal MPAs, for compliance monitoring. Real-time vessel tracking from AIS data supports enforcement by providing alerts of vessels crossing a virtual fence. For those vessels not emitting AIS signals, real-time detection is possible using directional arrays of hydrophones, over-the-horizon radar, and satellite-based detectors.
6.4 SCALABILITY
It is well accepted that oceanic biological processes are an important component in the global carbon cycle (Table 6.1). But perhaps the biggest question surrounding ecosystem protection and recovery regarding carbon removal is whether such conservation efforts will scale to the level of other proposed C removal efforts in the oceans and on land. To date, many of the studies on marine carbon have been conducted on relatively small geographic and temporal scales, although there have been several efforts to examine global C budgets for macroalgae, benthic algae, marine sedimentary carbon, the mesopelagic pump, and whale and fish carbon. The opportunistic nature of several of these studies and a reliance on intricate food web models compounded by a scarcity of observations to constrain models on a global scale can result in large uncertainties (Bianchi et al., 2021). Below, we briefly discuss questions of geographic and temporal scalability.
Geographic Scalability
The restoration of marine ecosystems could play an important role in the removal of carbon from the atmosphere, but to our knowledge, there have been no models addressing scalability on an ocean-basin or ecosystem scale that cut across species and processes. There is a need to develop an ecosystem-level approach to understanding the role of organisms and their ecosystem functioning in benthic, pelagic, tropical, temperate, and polar systems. New efforts to protect and restore the oceans, such as expanding MPAs and improving fisheries management, provide an ideal opportunity to study changes in C sequestration between degraded and restored ecosystems and between depleted and abundant marine species.
Marine ecosystems are diverse, and distributions and productivity are patchy. Continental shelves, for example, have been argued to contribute disproportionately to the oceanic uptake of CO2 (Fennel et al., 2018). There is a need to identify hot spots for C storage and sequestration. Locating connected macroalgal–sediment systems, for example, will be an important step toward including them in management actions aimed at carbon removal (Queirós et al., 2019). In the case
of nutrient cycling by large vertebrates and other animals, models have been used to assess scale and the potential impact on the global carbon cycle, often focusing on the Southern Ocean or other high-nutrient, low-chlorophyll (HNLC) regions (e.g., Martin et al., 2021; see also the scalability section in Chapter 5). As with ocean iron fertilization (OIF), the upwelling of the nutrients and respired carbon trapped at depth in the Southern Ocean could offset a substantial fraction of the naturally mediated downward flux of POC (Jin et al., 2008).
Even if the scale of some of these systems is relatively small, there could be an opportunity for island nations with limited land cover but large ocean Exclusive Economic Zones (EEZs) to offset their carbon emissions by protection efforts that explicitly value marine organisms and ecosystems in their waters. Similarly, the positive consequences of population, species, and habitat recovery and restoration could be substantial for ocean resilience and biodiversity on a global scale.
Temporal Scalability
Marine ecosystems have the potential to store carbon for long periods of time. Yet given current challenges, such as overfishing, pollutants, and climate change, what is the likelihood that they can be restored on a policy-relevant timescale? Duarte et al. (2020) suggest that substantial recovery of the abundance, function, and structure of marine life could be achieved by 2050 if major pressures are reduced. Response to protection efforts will vary by taxa and ecosystem, and some recovery efforts, such as the protection of coral reefs and endangered cetaceans such as North Atlantic and North Pacific right whales, will require substantial policy changes and sharp reduction in greenhouse gases.
Just as there are hot spots of C sequestration, there are likely hot moments, with sequestration rates varying seasonally and annually. As C emissions approach net zero, a diverse portfolio of marine conservation efforts may have the potential to contribute to global CDR approaches. (See the section on Durability for further discussion of temporal aspects of C sequestration under current and future scenarios.)
6.5 VIABILITY AND BARRIERS
Many people have strong emotional connections to oceans (Spence et al., 2018). These connections can result in greater resistance to engineered solutions and greater attraction to ecosystem-based solutions that endorse healthy and productive oceans while protecting marine wilderness (Cox et al., 2020). There is some support among the environmental NGO community for CDR efforts that involve ecosystem recovery on land and in the ocean. Social science research has indicated that ecosystem-based C removal efforts have greater public support than engineered approaches (Wolske et al., 2019). There is real value in conservation, yet the question of scale of ecosystem protection—how much carbon can be stored and removed by conservation and restoration efforts—is an area of active debate, one that is ripe for research.
Widespread efforts to protect and restore ecosystems could face barriers, because marine conservation can come into conflict with other uses, such as commercial fisheries, shipping, marine renewable energy, and mining. The designation of MPAs, for example, has faced opposition from fisheries and other industries. Conversely, the designation of large MPAs could reduce fossil fuel extraction, but also impede the expansion of renewables such as offshore wind. If the goal of establishing an MPA includes CDR, then managers should be open and honest about the objectives, uncertainties, and need for monitoring. Carbon benefits are just one of many ecosystem services that MPAs can provide, and they might need to be balanced against other objectives (Howard et al., 2017).
Research on MPAs has documented how MPA development and stricter fisheries management can impact local ocean-dependent communities and island nations. MPAs that do not incorporate
local interests and priorities can cause impacts on food security, displacement, and economic strain. Although MPAs can enhance food security, impacts vary among social groups (Mascia et al., 2010), and strategies must take this into account. Local perspectives are critical to understanding social impacts (Gollan and Barclay, 2020), and participatory approaches are one way to take small-scale fisheries into account (Kockel et al., 2020).
Any attempt to use ecosystem protection and recovery for C removal would likely benefit from the four guidelines proposed by a large community of researchers and conservation practitioners based in the UK for nature-based solutions to climate change: (1) ecosystem-based C removal must not be an alternative to the rapid phaseout of fossil fuels and must not delay decarbonization, (2) it must include a diversity of landscapes and in this case marine ecosystems, (3) it must be employed with the full engagement and consent of local communities and Indigenous peoples and promote adaptive capacity, and (4) it must be designed to provide quantifiable benefits for biodiversity (Seddon et al., 2021).
Environmental Impact
Despite the many uncertainties surrounding ecological restoration as a CDR tool, a clear advantage is that the environmental impact is almost certain to be positive, at least from the perspective of environmental stewardship and ecosystem integrity. There is also high economic value in marine stewardship (Roman et al., 2018). The environmental impact of any research agenda, deployment of monitoring equipment or research cruises, is likely to be small compared to C offsets or other more active CDR approaches.
In establishing a research agenda, it is important to acknowledge that we are not certain that there will be major C benefits, but we need to understand mechanisms and cycles at play. Such an approach could attract strong public support.
Co-benefits
Unlike some of the other ocean-based approaches considered in this report, there are considerable co-benefits in restoring ocean ecosystems, including biodiversity conservation and the restoration of many ecological functions and ecosystem services damaged by human activities such as pollution and overfishing. The recovery of kelp forests, for example, has C, biodiversity, and fisheries benefits (Duarte et al., 2020). The restoration of many species and habitats would also likely be beneficial for tourism and recreational and subsistence fishing; by encouraging the support of thriving marine life and habitats it is even possible that such restoration could mitigate the climate impacts of recreation.
In addition to any C benefits, reversing trends of biodiversity loss also have important nonuse values such as existence and spiritual values. The loss of whales and whale-fall habitats as a result of commercial whaling, for example, likely caused the endangerment and extinction of species dependent on these lipid-rich carcasses in the deep sea (Smith et al., 2019b). In addition to protecting the whales themselves, the recovery of whale populations could have C benefits and help protect and restore deep-sea communities. In some cases, these co-benefits might be the primary advantage of ecosystem restoration, and the uncertainties surrounding the geographic and temporal scale of CDR suggest that research into the long-term viability of many ecosystem-based approaches is essential.
Cost
Relative to other interventions, ecosystem-based solutions have the potential to be cost-effective, providing many benefits beyond CDR (Dasgupta, 2021). Nonetheless, nature-based solutions have been undercapitalized, and this lack of finance has been one of the barriers to the implementation and monitoring of such projects (Seddon et al., 2020). Similar to coastal blue carbon initiatives, the costs of implementing ecosystem recovery efforts for CDR depend on project size, intervention type, and monitoring of C removal (NASEM, 2019). If the effort is initiated as part of broader policies such as the Marine Mammal Protection Act, 30x30,4 or international treaties, only the costs of measuring and monitoring CDR need to be considered, but these policy actions come with their own costs. Although many of these costs are likely to be borne by government as part of efforts to protect the ocean, there is increased interest among private financial investors to support restoration and conservation projects: approximately 3 percent of impact assets, or $3.2B, are associated with conservation, and interest is growing among investors (Bass et al., 2018; Dasgupta, 2021).
Note that although direct costs are relatively low for the implementation of many ecosystem-based natural climate solutions, there can be negative economic impacts to some industries, such as oil and gas, fisheries, and mining, that provide jobs and lease payments for use.
Energy
As is true with costs, under most of the scenarios presented here, the energy required for research activities would be restricted to measuring and monitoring CDR associated with ecosystem and species recovery. These studies would likely be focused on research cruises and equipment necessary to measure marine biogeochemistry and changes in the carbon cycle.
Governance
Conservation efforts, such as eliminating overexploitation, protecting habitats, and restoring extirpated species, are generally feasible, local, and have high global governability (Gattuso et al., 2018). In addition to the co-benefits and public acceptance of ecosystem-based approaches, one of the great advantages of focusing on the protection and restoration of marine habitats and species is that the goals tend to align with those of several broad-reaching international agreements and domestic laws.
Chapter 2 provides an overview of the current state of the legal framework for ocean CDR. Many of the international and domestic laws discussed in that chapter could apply to the ecosystem recovery approaches discussed in this chapter. Several international agreements recognize the importance of protecting marine species and their habitats. Most notably:
- Parties to the United Nations Convention on the Law of the Sea (UNCLOS) have an “obligation to protect and preserve the marine environment” and must, among other things, take steps to prevent pollution and other activities that “may cause significant and harmful changes” to the marine environment.5 UNCLOS further provides that, whereas parties have a “sovereign right to exploit their natural resources,”6 they must adopt “proper conservation and management measures” to ensure that living resources within their EEZs
___________________
4 See https://www.oceanunite.org/30-x-30/.
5 Art. 192, 194, & 196, United Nations Convention on the Law of the Sea, 1833 U.N.T.S. 397, Dec. 10, 1982 (hereinafter “UNCLOS”).
6 Art. 193, UNCLOS.
- are “not endangered by over-exploitation.”7 Parties must also take steps to conserve the “living resources of the high seas.”8
- The Convention on Biological Diversity (CBD) requires parties to “[d]evelop national strategies, plans or programmes for the conservation . . . of biological diversity.”9 To that end, parties must “[e]stablish a system of protected areas or areas where special measures need to be taken to conserve biological diversity” and take steps to “restore degraded ecosystems and promote the recovery of threatened species.”10
A new international agreement, aimed at conserving marine biodiversity in areas beyond national jurisdiction, is also being developed by the UN General Assembly.
Many U.S. laws similarly recognize the importance of protecting marine ecosystems; for example:
- The Endangered Species Act (ESA) notes that many marine, freshwater, and terrestrial species have become extinct or at risk of extinction “as a consequence of economic growth and development untempered by adequate concern and conservation” (16 U.S.C. § 1531(a)). The ESA aims to reverse this trend by establishing a program for “the conservation of . . . endangered and threatened species” and “the ecosystems upon which they depend” (16 U.S.C. § 1521(b)).
- The Marine Mammal Protection Act (MMPA) similarly recognizes that certain “marine mammals are, or may be, in danger of extinction or depletion as a result of man’s activities” (16 U.S.C. § 1361(1)). The MMPA further declares that such species are an important resource that “should be protected” so as to “maintain the health and stability of marine ecosystems” (16 U.S.C. § 1361(6)). The MMPA aims to maintain stocks of marine mammals at their optimum sustainable populations, such that they continue to be “significant functioning element[s]” of their ecosystems (16 U.S.C. § 1461(2)).
- The National Marine Sanctuaries Act (NMSA) indicates that “certain areas of the marine environment” have “special national, and in some cases international, significance” and thus require special protection (16 U.S.C. § 1431(a)). The NMSA establishes a program through which such areas are identified and conserved (16 U.S.C. § 1431(b)(1)-(2)). A key goal of the program is to “maintain the natural biological communities in the national marine sanctuaries and to protect . . . natural habitats, populations, and ecological processes” (16 U.S.C. § 1431(b)(3)).
The ecosystem recovery approaches proposed in this chapter are broadly consistent with the above goals. They are also in line with the goal set by President Biden in January 2021 of conserving at least 30 percent of U.S. land and waters by 2030.11 A preliminary report on pathways for achieving the goal was published by the Departments of Agriculture, Commerce, and Interior and the Council on Environmental Quality (National Climate Task Force, 2021). The report emphasizes the importance of restoring ocean ecosystems, while also recognizing the “need to fight climate change with the natural solutions that . . . the oceans provide” (National Climate Task Force, 2021, p. 6). The conservation efforts discussed in this chapter are able to do both.
___________________
7 Art. 61, UNCLOS.
8 Art. 117, UNCLOS.
9 Art. 6(a), Convention on Biological Diversity, June 5, 1992, 1760 U.N.T.S. 79, June 5, 1992 (hereinafter CBD). 10 Art. 8, CBD.
11 Executive Order 14008 of January 27, 2021: Tackling the Climate Crisis at Home and Abroad, 86 Fed. Reg. 7619, 7627 (Feb. 1, 2021).
6.6 SUMMARY OF CARBON DIOXIDE REMOVAL POTENTIAL
The criteria for assessing the potential for ecosystem recover as a feasible approach to ocean CDR, described in Sections 6.2–6.5, is summarized in Table 6.3. The assessment presented is considered conservative and may change with future research; for example, future research may indicate that upper limits are higher for both scalability and durability.
6.7 RESEARCH AGENDA
We developed a research agenda (Table 6.4) that examines the potential role of protecting and restoring marine ecosystems in reducing additional C emissions and enhancing CDR. This agenda includes basic scientific research on the C removal potential and permanence of different organisms, ecosystems, and processes, an examination of the expected outcomes from different policy tools, and the socioeconomic and governance aspects of managing marine ecosystems and organisms for C removal. The framework for these studies combines experimental research, demonstration sites, and adaptive management.
For each of these systems, there is a need for research to help understand historical baselines in the context of the carbon cycle, past habitat degradation and organism losses due to human activities, and the potential changes to the characteristics of these systems as a result of climate perturbation and ocean acidification. It would be helpful to estimate the current and historical contribution of different species across the entire size spectrum in terms of annual C flux. Although there have been some efforts to estimate the CDR potential of marine recovery, such as changes in fisheries management and kelp afforestation, to our knowledge, there have been no broad solicitations for proposals at the federal level to address this nascent field.
Research will be required to follow the changes in C storage due to CDR as a result of management interventions such as the restoration of macroalgal habitats; the management of fish, whale, and other animal populations; and the establishment of MPAs. How much carbon is stored if ecosystems are restored? How much carbon is stored indirectly? These questions require more study. Given the somewhat piecemeal approach to studies of the coupling of oceanic ecosystems and carbon to date, there is a strong need for an ecosystems approach that combines both population dynamics, nutrient ecology and subsidies, and anthropogenic impacts. The framework developed for this study should be an integral part of long-term ecosystem-based management for the oceans, including CDR, fisheries, and other human uses.
This research agenda will likely require the development or use of current observing systems that can detect a change in C sequestration over a relatively short period of time (e.g., 10 years). A modeling effort will almost certainly be necessary to understand the C impacts of improved habitat and recovered populations over a measurable area. Indirect evidence from remineralized carbon and nutrients or associated oxygen consumption at the seafloor could help understand the role of animals and other marine organisms in these processes. For all of the approaches below, it will be essential to quantify the balance between C storage, burial, and release. It will also be important to show changes in C accumulation over time, both in the short term and the expected retention over decades and even centuries.
These efforts will require interdisciplinary work: the long separation of ecosystem ecology, population biology, and oceanography must end and go broader, expanding into economics, governance, and social science. Although each of the approaches below is not detail, it is expected that a full rollout would require several steps, including basic research, development, demonstration, deployment, and monitoring and verification (see NASEM, 2019). Key to the development of any place-based project is understanding the location and areal extent of the species or habitats considered below. More work is needed to develop remote sensing approaches to estimate potential areas
TABLE 6.3 CDR Potential of Ecosystem Restoration Efforts
| Knowledge base What is known about the system (low, mostly theoretical, few in situ experiments; medium, lab and some fieldwork, few carbon dioxide removal (CDR) publications; high, multiple in situ studies, growing body of literature) |
Low–Medium There is abundant evidence that marine ecosystems can uptake large amounts of carbon and that anthropogenic impacts are widespread, but quantifying the collective impact of these changes and the CDR benefits of reversing them is complex and difficult. |
| Efficacy What is the confidence level that this approach will remove atmospheric CO2 and lead to net increase in ocean carbon storage (low, medium, high) |
Low–Medium Confidence Given the diversity of approaches and ecosystems, CDR efficacy is likely to vary considerably. Kelp forest restoration, marine protected areas, fisheries management, and restoring marine vertebrate carbon are promising tools. |
| Durability Will it remove CO2 durably away from surface ocean and atmosphere (low, <10 years; medium, >10 years and <100 years; high, >100 years), and what is the confidence (low, medium, high) |
Medium 10–100 years The durability of ecosystem recovery ranges from biomass in macroalgae to deep-sea whale falls expected to last >100 years. |
| Scalability What is the potential scalability at some future date with global-scale implementation (low, <0.1 Gt CO2/yr; medium, >0.1 Gt CO2/yr and <1.0 Gt CO2/yr; high, >1.0 Gt CO2/yr), and what is the confidence level (low, medium, high) |
Low–Medium Potential C removal <0.1–1.0 Gt CO2/yr (low–medium confidence) Given the widespread degradation of much of the coastal ocean, there are plenty of opportunities to restore ecosystems and depleted species. However, ecosystems and trophic interactions are complex and changing, and research will be necessary to explore upper limits. |
| Environmental risk Intended and unintended undesirable consequences at scale (unknown, low, medium, high), and what is the confidence level (low, medium, high) |
Low (medium–high confidence) Environmental impacts would be generally viewed as positive. Restoration efforts are intended to provide measurable benefits to biodiversity across a diversity of marine ecosystems and taxa. |
| Social considerations Encompass use conflicts, governance-readiness, opportunities for livelihoods, etc. |
Trade-offs in marine uses to enhance ecosystem protection and recovery. Social and governance challenges may be less significant than with other approaches. |
| Co-benefits How significant are the co-benefits as compared to the main goal of CDR and how confident is that assessment |
High (medium–high confidence) Enhanced biodiversity conservation and the restoration of many ecological functions and ecosystem services damaged by human activities. Existence, spiritual, and other nonuse values. Potential to enhance marine stewardship and tourism. |
| Cost of scale-up Estimated costs in dollars per Mt CO2 for future deployment at scale; does not include all of monitoring and verification costs needed for smaller deployments during R&D phases (low, <$50/t CO2; medium, ~$100/t CO2; high, >>$150/t CO2) and confidence in estimate (low, medium, high) |
Low <$50/t CO2 (medium confidence) Varies, but direct costs would largely be for management and opportunity costs for restricting uses of marine species and the environment. No direct energy used. |
| Cost and challenges of carbon accounting Relative cost and scientific challenge associated with transparent and quantifiable carbon tracking (low, medium, high) |
High Monitoring net effect on C sequestration is challenging. |
| Cost of environmental monitoring Need to track impacts beyond carbon cycle on marine ecosystems (low, medium, high) |
Medium (medium–high confidence) All CDR will require monitoring for intended and unintended consequences both locally and downstream of CDR site, and these monitoring costs may be substantial fraction of overall costs during R&D and demonstration-scale field projects. This cost of monitoring for ecosystem recovery may be lower. |
| Additional resources needed Relative low, medium, high to primary costs of scale-up |
Low Most recovery efforts will likely require few materials and little energy, though enforcement could be an issue. Active restoration of kelp and other ecosystems would require more resources. |
for restoration and protection for macroalgae and benthic algae, as well as marine animal carbon, with a deeper understanding of historic biomass and expectations for populations in a future altered by climate change and habitat modifications.
In addition to determining the C sequestration potential of ecosystem restoration, this work is expected to make important contributions to marine ecology. Uniting researchers in ecosystem ecology, biological oceanography, population biology, and social science—disciplines that have long operated on separate paths—to examine marine CDR can provide co-benefits by coordinating research, analyzing and synthesizing information, and drafting and implementing management plans.
Habitat Protection and Restoration
The current push to increase MPAs provides a unique opportunity to examine the impacts of marine conservation and restoration on CDR. Policies such as 30 x 30, calling for 30 percent protection of the oceans in less than a decade, could provide research platforms to measure changes in C uptake and sequestration in relation to macroalgae, marine animal carbon, benthic algae, and sedimentary carbon. By protecting multiple sites, there will be opportunities to compare ecosystem functions across protected areas as well as inside and outside of protected areas. These could serve as experimental treatments that can be leveraged to test ideas about ocean CDR linked to ecosystems. As with coastal blue carbon systems, assessing the C stocks of offshore MPAs, and their changes over time, will require mapping, sample collection and analysis, monitoring C movement into and out of the system, and determining emissions avoided because of management activities (Howard et al., 2017). One potentially effective approach to understanding the CDR potential of restored ecosystems is to compare the present-day C fluxes in the ocean to preindustrial C transfer. By quantifying the human perturbations of C flux in the ocean, we can estimate the potential role of a restored ocean in the carbon cycle (see, e.g., Regnier et al., 2013).
Historically, much of the ocean economy has been based on commercial extraction, such as hunting fisheries, whaling, and guano, though this has been changing in recent years (e.g., Roman et al., 2018). A shift away from the overharvest of marine resources in the twentieth century is likely to result in changes in the carbon cycle: by reduction in the extraction of fish biomass, fuel consumption, and habitat disturbance and through changes in trophic structure. There have been many calls to halt the loss of biodiversity as fast as possible and to reach net positive results through restoration and regeneration of marine and terrestrial ecosystems (e.g., naturepositive.org). We strongly recommend that C accounting and research be a central aspect of the monitoring of MPAs
and restoration efforts in the coming decades, including the examination of multiple systems with geographic, ecological, and taxonomic differences and varying levels of anthropogenic pressure. There is a need to develop consistent methodologies to conduct economic analyses and examine trade-offs between C benefits and other ecosystem services (e.g., Boumans et al., 2015).
Community engagement is essential here. Social science research can help us understand the institutions, policies, and cultural practices that lead to community support and engagement in marine ecosystem recovery. It can also help determine why efforts sometimes fail and what can make them successful. A governance component could examine how the legal framework for MPAs could be structured to increase C benefits. Since almost 50 percent of sedimentary C stocks reside outside of EEZs, international cooperation will be required to prevent remineralization of C stored beneath the high seas (Atwood et al., 2020). Ultimately, a transdisciplinary research program that includes questions of governance, oceanographic, conservation, and ecological research activities, and stakeholder engagement is likely to result in the greatest success.
Macroalgae
Research is needed to address numerous ecological, social, and biogeochemical questions in relation to the protection and restoration of macroalgal communities. It is essential to quantify the balance between C burial and release. What is the permanence of this buried material, and how vulnerable is it to rising temperatures and elevated climate change disturbances? Measurements of detritus production by whole macroalgal communities are needed to estimate how much released macroalgal particulate organic carbon is likely to become sequestered in deep, coastal sediment or other habitats (Queirós et al., 2019). Sequestration rates could be small if much of the POC is consumed and respired back to the surface ocean or if it settles on the seabed along the continental shelf. What is the natural mechanism for transporting POC away from the coast and then downward into the deep ocean? Stable isotopes and eDNA could be employed more widely to quantify and trace the contribution of macroalgae to deep-sea sediments, both in the past and present.
There is also a need for improved estimates of the global area and production of macroalgae at the level of major functional groups (Krause-Jensen et al., 2018). There has been some progress on this front using high-resolution satellite imagery, such as efforts to map the distribution and persistence of giant kelp (Macrocystis pyrifera) forests across a 35-year time series (Arafeh-Dalmau et al., 2021). Such techniques could be applied to monitor kelp protection and restoration efforts. Modeling frameworks would also be essential to understand the spatial distribution of macroalgal biomass and its contribution to climate budgets and C sequestration (van Son et al., 2020). There is also a need to better understand the impacts of elevated temperatures and other climate impacts on macroalgal distribution and C storage.
Natural and social science research on the actions that could be used for expanding natural macroalgae is needed, including studying the costs and benefits of deployment and if the actions are effective. Kelp restoration is becoming a prominent management intervention, but it can be quite expensive (more than $6,000 per hectare; Eger et al., 2020). Studying restoration success at appropriate spatial scales will be essential to encourage institutions to support kelp restoration.
Benthic Communities
There is a need to extend our understanding of the contributions that benthic communities make in relation to greenhouse gases, C sequestration and storage, and other climate change–related measures (Solan et al., 2020). Although there has been a substantial amount of work on shallow and intertidal benthic systems, such as mangroves and seagrasses, offshore and deep-sea benthic communities have largely been overlooked. Even shallow-water benthic communities, such as
oyster reefs and other bivalve populations, could benefit from increased scrutiny: calcium carbonate (CaCO3) shell production decreases alkalinity and can act as a source for CO2, yet shellfish reefs contain significant pools of carbon and facilitate atmospheric C drawdown via filtration and biodeposition (Fodrie et al., 2017; Solan et al., 2020). Understanding the balance between these processes is essential to quantify the C dynamics of benthic communities.
A research agenda that explores benthic protection and restoration would expand our understanding of these and surrounding areas. Benthic algae rarely occur in isolation: in designing a research agenda, it is important to move beyond individual species to consider ecosystem-level responses. Approaches should measure the balance of C release and burial, or net community calcification versus photosynthesis, at the ecosystem level.
What is the extent of C removal at the ecosystem level as a result of human activities such as trawling and recreation? How can these activities be mitigated? An interdisciplinary approach that examines how these changes could affect key stakeholders is more likely to succeed from a carbon, conservation, and societal perspective.
Marine Animals and Carbon Dioxide Removal
Given the many uncertainties surrounding CDR and marine animals, quantifying the direct and indirect roles of marine fauna in the carbon cycle is a priority for further research (Stafford et al., 2021). Synthesis studies to identify the potential significance of marine vertebrates in regional and ocean basin scales is essential, to see whether their functional impacts are greater than the relatively small biomass (compared to phyto- and zooplankton) and compare them to other carbon cycle components (Martin et al., 2021).
Of perhaps equal value is examining the role of fisheries and other human activities, such as shipping, on marine populations and how the mitigation of some of the resulting environmental impacts could affect CDR. Essential in these studies is understanding the approaches necessary to ensure that reducing an activity such as bottom trawling in one area does not increase trawling in another, resulting in no net gain for carbon. An international and multistakeholder approach will likely be necessary here. A governance component of this research could examine how the legal framework for protecting marine mammals, seabirds, fish, and other large animals could be structured to increase C benefits.
A recent report and workshop at the International Whaling Commission identified data gaps and research needs on the role of whales in the carbon cycle (Roman et al., 2021). Although several studies have examined the ecological role of cetaceans, understanding these mechanisms, especially regarding nutrient transport and cycling, in the context of CDR is still in its infancy. Empirical investigations and additional study sites are needed to enhance, support, or refute hypotheses regarding the role of cetaceans in the carbon cycle. Research on the vulnerability of different species and populations to rising sea surface temperatures and reduced sea ice will help determine which species and populations, along with their CDR potential, are likely to be affected by climate change in the next century (e.g., van Weelden et al., 2021).
Marine Biomass and Deadfall Carbon
The CDR potential of pelagic communities has not been as well examined as coastal and nearshore benthic systems. The development stage of research for the restoration of marine animals involves understanding historic numbers and ranges, but also projecting changes with altered habitats and climates. As biomass and population numbers change, it will be essential to look for signals, via changes in organic and inorganic C stocks, nutrients, or associated oxygen consumption, in the water column and at the seafloor, on both local and larger scales. In addition to changes
prompted by conservation, there is a need for further analyses of shifts in species habitat and the potential influence on food webs and the carbon cycle. Understanding the impacts of the expansion of salps into former krill ranges in the Antarctic and the potential biogeochemical changes, for example, will help track the role of animals in the carbon cycle now and in the future ocean (Böckmann et al., 2021).
Among cetaceans, generalist feeders such as humpback whales appear to be thriving under protection regimes. Other species such as the North Atlantic right whale have not responded to restrictions on direct harvest and remain endangered. There is an urgent need to examine past population sizes, current trends in marine vertebrate populations, and expectations for species and ecological processes under climate change scenarios. With data on life history and population structure, it is possible to quantify the amount of carbon stored in living biomass and sequestered in deadfalls. The most robust quantification currently available was based on global population estimates from 2001 (Pershing et al., 2010). This analysis could be updated with new population estimates and better understanding of pre-exploitation and future numbers.
Nutrient Cycling
The role of iron and nitrogen in the ocean carbon cycle has been well established, yet much of the focus on nutrient fertilization has been on open-ocean iron addition experiments and a few macronutrient (nitrogen and phosphorus) studies or schemes (see Chapter 3). There is a tendency to favor more natural approaches over engineered or artificial approaches; this preference could result in a dilemma between approaches with high C sequestration potential but low levels of acceptability and approaches with possibly low sequestration potential but high levels of acceptability (Bertram and Merk, 2020). At present, we do not know the sequestration potential of restoring historic populations of marine animals such as whales and seabirds or of shifting regimes under climate change, such as jellification, including salps.
There is a strong need for empirical data to test the role of nutrient cycling in the ocean carbon cycle. Although several models have been developed to look at the role of fecal nitrogen and iron in primary productivity (e.g., Lavery et al., 2010; Roman et al., 2014; Ratnarajah et al., 2016), it is unknown how closely these results approximate field conditions. Further quantification of cetacean diving behavior is needed to determine what proportion of foraging dives occur above or below the mixed layer and thus contribute to autochthonous or allochthonous nutrient cycling. It is also important to determine how available fecal plume nutrients are to phytoplankton and the impact of the microbial loop on nutrient uptake (Ratnarajah et al., 2018). Finally, it is critical to estimate what proportion of the carbon fixed as a result of whale fecal fertilization sinks below the mixed layer and becomes stored or sequestered (Roman et al., 2021). A spatial understanding of sequestration timescales is important here. For many areas of the ocean, it is likely that upper-ocean ecosystem productivity will be transported back to the surface within 50 years, and thus will have only a short-term influence on atmospheric CO2 (Siegel et al., 2021a).
Predator and Prey Dynamics
The loss of apex predators in ocean ecosystems has resulted in changes in marine food webs, ecosystems, and productivity. The restoration of these and other large animals has the potential to increase C sequestration (e.g., the trophic interactions between sea otters, sea urchins, and kelp in the North Pacific) or decrease C storage (green sea turtles feeding on seagrasses in the Indian Ocean). The examination of changes in C dynamics as these predator–prey dynamics are restored offers a unique opportunity for marine ecologists, population biologists, oceanographers, and social scientists to work together. They can help develop the tools necessary to prompt these changes and
examine the shifts in biomass, ecological functions, ecosystem services, and C sequestration over time.
6.8 SUMMARY
Efforts to protect and restore marine ecosystems cannot be considered in isolation from the challenge of reducing the production of greenhouse gases. These goals of restoring habitats and recovering overharvested species are ambitious but necessary and could help reduce atmospheric carbon in coming decades, achieving other co-benefits along the way (Hoegh-Guldberg et al., 2019). This chapter identifies some of the approaches for marine ecosystem restoration that could contribute and the processes by which this restoration contributes to C removal, going beyond the current baseline of sequestration.
Most of the signatories of the Paris Agreement have committed to nature-based solutions in their climate change programs (Seddon et al., 2019). A whole ecosystem perspective, which includes abundant species and protected ecosystems as well as those in need of restoration, will be essential to these approaches, along with better understanding of the ecological functions provided by marine species. Leveraging naturally occurring C fixation, storage, and sequestration interactions in the oceans and on land, can be a strategy with lower risk and cost than many geoengineering solutions (Griscom et al., 2017; Martin et al., 2021).
Whether ecosystem restoration is scalable is a matter of perspective and timing. Although the CDR potential of ecosystem recovery in the oceans is relatively small in comparison to present greenhouse gas emissions, this is perhaps more a reflection of the magnitude of current global emissions than a diminished role in the seas. As emission rates come down, ocean CDR could rise in importance. In anticipation of that transition, ecosystem-based approaches merit discreet attention and funding now.
The challenge for ecosystem-based CDR does not center around functionality—clearly animals, plants, and microbes all play an important role in the carbon cycle of the ocean—rather it is a question of scaling, verification, and concerns about reversibility. Restoring and protecting the ocean will likely remove a substantial amount of carbon from the atmosphere (though even approximate estimates remain challenging at this point) with relatively low costs for CDR verification above those already allocated for conservation efforts. As C emissions approach net zero, a diverse portfolio of marine conservation efforts could make a substantial contribution to global CDR approaches.
TABLE 6.4 Research and Development Needs: Ecosystem Recovery
| # | Recommended Research | Gap Filled | Environmental Impact of Research | Estimated Cost of Research ($M/yr) | Time Frame (years) |
|---|---|---|---|---|---|
| 6.1 | Restoration ecology and carbon | Estimate the change in C storage between natural and present-day marine ecosystems | Low | 8 | 5 |
| 6.2 | Marine protected areas (MPA): Do ecosystem-level protection and restoration scale for marine CDR? | Estimate the ability of ocean conservation and MPA protection to enhance the storage and sequestration of carbon per year until 2050 | Low | 8 | 10 |
| 6.3 | Macroalgae: Carbon measurements, global range, and levers of protection | Improve our understanding of the fate of macroalgal carbon, the range of different species and habitats, and the socioeconomic levers and costs of restoring kelp and other macroalgal habitats | Low | 5 | 10 |
| 6.4 | Benthic communities: disturbance and restoration | Improve our understanding of the impacts of human disturbance on benthic communities and the potential rate of change under different protection scenarios | Low | 5 | 5 |
| 6.5 | Marine animals and CDR | Carbon interactions and outcomes have been estimated for only a few marine species. Efforts will improve our understanding of the direct and indirect impacts of marine animals on CDR, including biomass, deadfall carbon, nutrient transfer, and trophic cascades | Low | 5 | 10 |
| 6.6 | Animal nutrient-cycling | Test of models of movement of iron (vertical) and nitrogen (horizontal) as mediated by whales and other air-breathing vertebrates | Low | 5 | 5 |
| 6.7 | Commercial fisheries and marine carbon | Improve our understanding of fisheries emissions, fish populations and ecological function, and impacts on sedimentary carbon | Low | 5 | 5 |
| 6.8 | Community engagement with marine ecosystem recovery | Understand what institutions, policies, and cultural practices lead to community support and engagement in marine ecosystem recovery; why efforts might fail; and what can make them successful | Low | 2 | 5 |
NOTE: Bold type identifies priorities for taking the next step to advance understanding of ecosystem recovery as a CDR approach.


































